

These little essays originally appeared as articles in ‘Longman’s Magazine,’ the ‘Cornhill Magazine,’ ‘Macmillan’s Magazine,’ the ‘Gentleman’s Magazine,’ and ‘Belgravia,’ and I have to thank the editors and proprietors of those periodicals for kind permission to reprint them here. They are now offered to the public as a first instalment of a work which I hope some day more fully to carry out—a Functional Companion to the British Flora. We know by this time pretty well what our English wild flowers are like: we want to know next why they are just what they are, and how they came to be so.
G. A.
Lyme, Dorset:
July 1883.
| PAGE | |||
| INTRODUCTORY | 1 | ||
| I. | THE DAISY’S PEDIGREE | 6 | |
| II. | THE ROMANCE OF A WAYSIDE WEED | 44 | |
| III. | STRAWBERRIES | 80 | |
| IV. | CLEAVERS | 100 | |
| V. | THE ORIGIN OF WHEAT | 133 | |
| VI. | A MOUNTAIN TULIP | 173 | |
| VII. | A FAMILY HISTORY | 197 | |
| VIII. | CUCKOO-PINT | 236 |
Our beautiful green England is carpeted, more than any other country in the world, perhaps, save only Switzerland and a few other mountain lands, with a perpetual sward of vivid verdure, interspersed with innumerable colours of daisies, and buttercups, and meadow-sweet, and harebells, and broader patches of purple heather. It is usual to speak of tropical vegetation, indeed, with a certain forced ecstasy of language; but those who know the tropics best, know that, though you may find a few exceptionally large and brilliant blossoms here and there under the breadth and shade of equatorial forests, the prevailing tone is one of monotonous dry greenery; and there is nothing anywhere in very southern climes to compare, as to mass of colour, with our Scotch hill-sides, our English gorse-clad commons, or our beautiful dappled meadows and[2] cornfields, all aglow with the infinite wealth of poppies, bluebottles, foxgloves, ox-eye daisies, and purple fritillaries. The Alps alone can equal the brilliant colouring of our own native British flora. Poor as it is in number of species—a mere isolated fragment of the wider European groups—it can fearlessly challenge the rest of the whole world in general mingled effect of gaiety and luxuriance.
Now, every one of these English plants and weeds has a long and eventful story of its own. In the days before the illuminating doctrine of evolution had been preached, all we could say about them was that they possessed such and such a shape, and size, and colour: and if we had been asked why they were not rounder or bigger or bluer than they actually are, we could have given no sufficient reason, except that they were made so. But since the great principle of descent with modification has reduced the science of life from chaos to rational order, we are able to do much more than that. We can now answer confidently: Such and such a plant is what it is in virtue of such and such ancestral conditions, and it has been altered thus and thus by these and those variations in habit or environment. Every plant or animal, therefore, becomes for us a puzzle to be explained, a problem to be solved, a hieroglyphic inscription to be carefully[3] deciphered. In the following pages, I have taken some half-dozen of familiar English weeds or flowers, and tried thus to make them yield up the secret of their own origin. Each of them is ultimately descended from the common central ancestor of the entire flowering group of plants; and each of them has acquired every new diversity of structure or appearance for some definite and useful purpose. As a rule, traces of all the various stages through which every species has passed are still visibly imprinted upon the very face of the existing forms: and one only requires a little care and ingenuity, a little use of comparison and analogy, to unravel by their own aid the story of their own remoter pedigree. This is the method which I have here followed in the papers that deal with the various modifications of the daisy, of the grasses, of the lilies, of the strawberry, and of the whole rose family.
Again, not only has each English plant a general history as a species, but it has also a separate history as a member of the British flora. Besides the question how any particular flower or fruit came to exist at all, we have to account for the question how it came to exist here and now in this, that, or the other part of the British Islands. For, of course, all plants are not to be found in all parts of the world alike, and their[4] distribution over its surface has to be explained on historical grounds just as a future ethnologist would have to explain the occurrence of isolated French communities in Lower Canada and Mauritius, of African negroes in Jamaica and Brazil, or of Chinese coolies in San Francisco and the Australian colonies. In this respect, our English plants open out a series of interesting problems for the botanical researcher; because we happen to possess a very mixed and fragmentary flora, made up to a great extent of waifs and strays from at least three large distinct continental groups, besides several casual colonists. Thus while at Killarney we get a few rare Spanish or Portuguese types, in Caithness and the Highlands we get a few rare Alpine or Arctic types: and while in Norfolk and Suffolk we find some central European stragglers, the ponds of the Hebrides are actually occupied by at least one American pond-weed, its seeds having been wafted over by westerly breezes, or carried unconsciously by water-birds in the mud and ooze which clung accidentally to their webbed feet. Moreover, we know that at no very remote period, geologically speaking, Britain was covered by a single great sheet of glaciers, like that which now covers almost all Greenland: and we may therefore conclude with certainty that every plant at present in the country[5] has entered it from one quarter and another at a date posterior to that great lifeless epoch. This, then, gives rise to a second set of problems, the problems connected with the presence in England of certain stray local types, Alpine or Arctic, Southern or Transatlantic, European or Asiatic. Questions of this sort I have raised and endeavoured to answer with regard to two rare English plants in the papers on the hairy spurge and the mountain tulip.
In short, these little essays deal, first with the evolution of certain plant types in general; and secondly with their presence as naturalised citizens of our own restricted petty insular floral commonwealth.

Fig. 1.—The Common Daisy.
Have you ever paused for a moment to consider how much man loses for want of that microscopic eye upon whose absence complacent little Mr. Pope, after his optimistic fashion, was apparently inclined rather to congratulate his fellow-beings than otherwise? What[7] a wonderful world we should all live in if only we could see it as this little beetle here sees it, half buried as he is in a mighty forest of luxuriant tall green moss! Just fancy how grand and straight and majestic those slender sprays must look to him, with their waving, feathery branches spreading on every side, a thousand times more gracefully than the long boughs of the loveliest tropical palm trees on some wild Jamaican hill-side. How quaint the tall capsules must appear in his eyes—great yellow seed-vessels nearly as big as himself, with a conical, pink-edged hood, which pops off suddenly with a bang, and showers down monstrous nuts upon his head when he passes beneath. Gaze closely into the moss forest, as it grows here beside this smooth round stone where we are sitting, and imagine you can view it as the beetle views it. Put yourself in his place, and look up at it towering three hundred feet above your head, while you vainly strive to find your way among its matted underbrush and dense labyrinths of close-grown trunks. Then just look at the mighty monsters that people it. The little red spider, magnified to the size of a sheep, must be a gorgeous and strange-looking creature indeed, with his vivid crimson body and his mailed and jointed legs. Yonder neighbour beetle, regarded as an elephant, would seem a terrible[8] wild beast in all seriousness, with his solid coat of bronze-burnished armour, his huge hook-ringed antennæ, and his fearful branched horn, ten times more terrible than that of a furious rhinoceros charging madly through the African jungle. Why, if you will only throw yourself honestly into the situation, and realise that awful life-and-death struggle now going on between an ant and a May-fly before our very eyes, you will see that Livingstone, and Serpa Pinto, and Gordon Cumming are simply nowhere beside you: that even Jules Verne’s wildest story is comparatively tame and commonplace in the light of that marvellous miniature forest. Such a jumble of puzzle-monkeys, and bamboos, and palms, and banyan trees, and crags, and roots, and rivers, and precipices was never seen; inhabited by such a terrible and beautiful phantasmagoria of dragons, hippogriffs, unicorns, rocs, chimæras, serpents, and wyverns as no mediæval fancy ever invented, no Greek mythologist ever dreamt of, and no Arabian story-teller ever fabled. And yet, after all, to our clumsy big eyes, it is but a little patch of familiar English grass and mosses, crawled over by half a dozen sleepy slugs and long-legged spiders, and slimy earthworms.
Still, if you so throw yourself into the scene, you cannot avoid carrying your own individuality with[9] you into the beetle’s body. You fancy him admiring that fairy landscape as you would admire it were you in his place, provided always you felt yourself quite secure from the murderous jaws and hooked feet of some gigantic insect tiger lurking in the bristly thicket behind your back. But, as a matter of fact, I greatly doubt whether the beetle has much feeling for beauty of scenery. For a good many years past I have devoted a fair share of my time to studying, from such meagre hints as we possess, the psychology of insects: and on the whole I am inclined to think that, though their æsthetic tastes are comparatively high and well-developed, they are, as a rule, decidedly restricted in range. Beetles and butterflies only seem to admire two classes of visible objects—their own mates, and the flowers in which they find their food. They never show much sign of deliberate love for scenery generally or beautiful things in the abstract outside the limits of their own practical life. If this seems a narrow æsthetic platform for an intelligent butterfly, one must remember that our own country bumpkin has perhaps a still narrower one; for the only matter in which he seems to indulge in any distinct æsthetic preference, to exercise any active taste for beauty, is in the choice of his sweetheart, and even there he is not always conspicuous for the refinement[10] of his judgment. But there is a way in which one can really to some extent throw oneself into the mental attitude of a human being reduced in size so as to look at the moss-forest with the eye of a beetle, while retaining all the distinctive psychological traits of his advanced humanity: and that is by making himself a microscopic eye with the aid of a little pocket-lens. Even for those who do not want to use one scientifically, it opens a whole universe of new and delightful scenery in every tuft of grass and every tussock of wayside weeds; and by its aid I hope to show you this morning how far the eyes and æsthetic tastes of insects help us to account for the pedigree of our familiar childish friend, the daisy. No fairy tale was ever more marvellous, and yet certainly no fairy tale was ever half so true.
I propose then, to-day, to dissect one of these daisies with my little knife and glass, and unravel, if I can, the tangled skein of causes which have given it its present shape, and size, and colour, and arrangement. If you choose, you can each pick a daisy for yourselves, and pull it to pieces as I go along, to check off what I tell you; but if you are too lazy, or can’t find one within reach, it doesn’t much matter; for you can at least carry the picture of so common a flower well enough in your mind’s eye to follow what[11] I have to say without one: and that is all that is at all necessary for my present purpose.
The question as to how the daisy came to be what it is, is comparatively a new one. Until a short time ago everybody took it for granted that daisies had always been daisies, cowslips always cowslips, and primroses always primroses. But those new and truer views of nature which we owe to Mr. Darwin and Mr. Herbert Spencer have lately taught us that every plant and every animal has a long history of its own, and that this history leads us on through a wonderful series of continuous metamorphoses compared with which Daphne’s or Arethusa’s were mere single episodes. The new biology shows us that every living thing has been slowly moulded into its existing shape by surrounding circumstances, and that it bears upon its very face a thousand traces of its earlier stages. It thus invests the veriest weed or the tiniest insect with a fresh and endless interest: it elevates them at once into complex puzzles for our ingenuity—problems quite as amusing and ten times as instructive as those for whose solution the weekly papers offer such attractive and unattainable prizes. What is the meaning of this little spur? How did it get that queer little point? Why has it developed those fluffy little hairs? These are the questions[12] which now crop up about every part of its form or structure. And just as surely as in surveying England we can set down Stonehenge and Avebury to its prehistoric inhabitants, Watling Street and the Roman Wall to its southern conquerors, Salisbury and Warwick to mediæval priests and soldiers, Liverpool and Manchester to modern coal and cotton—just so surely in surveying a flower or an insect can we set down each particular point to some special epoch in its ancestral development. This new view of nature invests every part of it with a charm and hidden meaning which very few among us have ever suspected before.
Pull your daisy to pieces carefully, and you will see that, instead of being a single flower, as we generally suppose at a rough glance, it is in reality a whole head of closely packed and very tiny flowers seated together upon a soft fleshy disk. Of these there are two kinds. The outer florets consist each of a single, long, white, pink-tipped ray, looking very much like a solitary petal: the inner ones consist each of a small, golden, bell-shaped blossom, with stamens and pistil in the centre, surrounded by a yellow corolla much like that of a Canterbury bell in shape, though differing greatly from it in size and colour. The daisy, in fact, is one of the great family[13] of Composites, all of which have their flowers clustered into similar dense heads simulating a single blossom, and of which the sunflower forms perhaps the best example, because its florets are quite large enough to be separately observed even by the most careless eye.
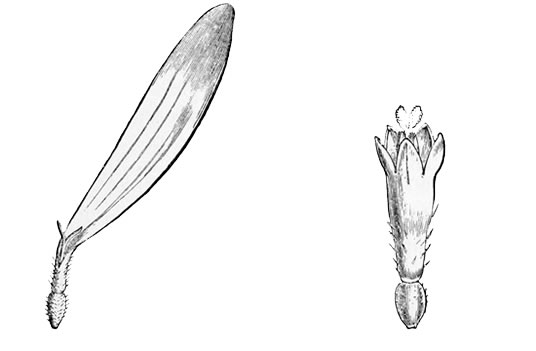
| Fig. 2.—Ray floret of Daisy. | Fig. 3.—Central floret of Daisy. |
Now, if you look closely at one of the central yellow florets in the daisy, you will see that its edge is vandyked into four or five separate pointed teeth exactly like those of the Canterbury bell. These teeth clearly point back to a time when the ancestors of the daisy had five separate petals on each flower, as a dog-rose or a May-blossom still has. Again,[14] before the flowers of the daisy had these five separate petals, they must have passed through a still earlier stage when they had no coloured petals at all. And as it is always simpler and easier to recount history in its natural order, from the first stages to the last, rather than to trace it backward from the last to the first, I shall make no apology for beginning the history of the daisy at the beginning, and pointing out as we go along the marks which each stage has left upon its present shape or its existing arrangement and colour.
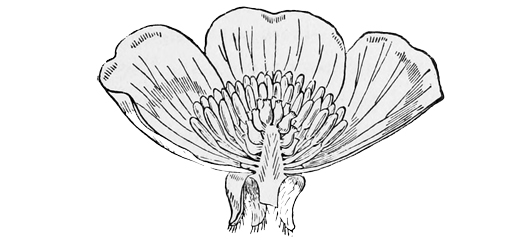
Fig. 4.—Longitudinal section of Common Buttercup.
The very earliest ancestor of the daisy, then, with which we need deal to-day, was an extremely simple and ancient flower, hardly recognisable as such to any save a botanical eye. And here I must begin, I fear, with a single paragraph of rather dull and technical matter, lest you should miss the meaning of some things I shall have to tell you in the sequel. If you look into the middle of a buttercup or a lily you know that you will see certain little yellow spikes and knobs within the petals, which form a sort of central rosette, and look as if they were put there merely to give finish and completeness to the whole blossom. But in reality these seemingly unimportant spikes and knobs are the most important parts, and the only indispensable parts, of the entire flower. The bright[15] petals, which alone are what we generally have in our minds when we think of flowers, are comparatively useless and inessential organs: a vast number of flowers have not got them at all, and, in those which have got them, their purpose is merely subsidiary and supplementary to that of the little central spikes and knobs. For the small yellow rosette consists of the stamens and pistils—the ‘essential floral organs,’ as botanists call them. A flower may be complete with only a single stamen or a single pistil, apart from any petals or other bright and conspicuous surroundings; and some of the simplest flowers do actually consist of such separate parts alone: but without stamens and pistils there can be no flower at all. The object of the flower, indeed, is to produce fruit and seed, and the pistil is the seed-vessel in its earliest form; while the stamen manufactures the pollen without which[16] the seeds cannot possibly be matured within the capsules. In some species the stamens and pistils occur in separate flowers, or even on separate plants; in others, the stamens and pistils occur on the same plant or in the same flower, and this last is the case in almost all the blossoms with which we are most familiar. But the fundamental fact to bear in mind is this—that the stamens and pistils are the real and essential parts of the flower, and that all the rest is leather and prunella—mere outer decoration of these invariable and necessary organs. The petals and other coloured adjuncts are, as I hope to show you, nothing more than the ornamental clothing of the true floral parts; the stamens and pistils are the living things which they clothe and adorn. Now probably you know all this already, exactly as the readers of the weekly reviews know by this time all about the personage whom we must not describe as Charlemagne, or the beings whom it is a mortal sin to designate as Anglo-Saxons. But then, just as there are possibly people in the worst part of the East End who still go hopelessly wrong about Karl and the Holy Roman Empire, and just as there are possibly people in remote country parishes who are still the miserable victims of the great Anglo-Saxon heresy, so, doubtless, there may yet be persons—say[17] in the western parts of Cornwall or the Isle of Skye—who do not know the real nature of flowers; and these persons must not be wholly contemned because they happen not to be so wise as we ourselves and the Saturday Review. An eminent statistician calculates that Mr. Freeman has demolished the truculent Anglo-Saxon in 970 several passages, and yet there are even now persons who go on firmly believing in that mythical being’s historical existence. And the moral of that is this, as the Duchess would say, that you should never blame any one for telling you something that you knew before; for it is better that ninety-nine wise men should be bored with a twice-told tale, than that one innocent person should be left in mortal error for lack of a short and not wholly unnecessary elementary explanation.
Fig. 5.—Frond and flower of Duckweed.
The simplest and earliest blossoms, then—to return from this didactic digression—were very small and inconspicuous flowers, consisting, probably, of a single stamen and a single pistil each. Of these simplest and earliest forms a few still luckily survive at the present day; for it is one of the rare happy chances in this queerly ordered universe of ours that evolution has almost always left all its footmarks behind it, visibly imprinted upon the earth through all its ages. When any one form develops slowly[18] into another, it does not generally happen that the parent form dies out altogether: on the contrary, it usually lingers on somewhere, in some obscure and unnoticed corner, till science at last comes upon it unawares, and fits it into its proper place in the scale of development. We have still several fish in the very act of changing into amphibians left in a few muddy tropical streams; and several oviparous creatures in the very act of changing into mammals left in the isolated continent of Australia; and so we have also many low, primitive, or simple forms of plants and animals left in many stray situations in every country. Amongst them are some of these earliest ancestral flowers. On almost every wayside pond you will find all the year round a green film of slimy duckweed. This duckweed is, as it were, the Platonic idea of a flowering plant—the generic type common to them all reduced to its simplest elements. It has no roots, no stem, no branches, no visible blossom, no apparent seed; it consists merely of solitary, roundish, floating leaves, budding out at the edge into other leaves, and so spreading till it covers the whole pond. But if you look closely into the slimy mass in summer time, you may be lucky enough to catch the weed in flower—though not unless you have a quick eye and a good pocket-lens.[19] The flowers consist of one, and sometimes two, stamens and a pistil, growing naked out of the edge of the leaf. No one but a botanist could ever recognise their nature at all, for they all look like mere yellowish specks on the slender side of the green frond; but the pistil contains true seeds, and the stamens produce true pollen, and from the botanical standpoint that settles the question of their floral nature at once. They are, in fact, representatives of the simplest original form of flower, preserved to our own day on small stagnant ponds, where the competition of other plants does not press them hard as it has pressed their congeners on dry land or in open[20] lakes and rivers.[1] From some such simple form as this we may be pretty sure that all existing flowering plants are ultimately descended.
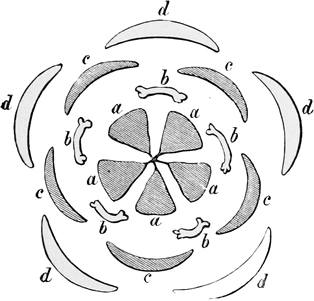
a, Carpels or ovaries; b, stamens; c, petals; d, calyx.
Fig. 6.—Diagram of primitive dicotyledonous flower.
In most modern flowers, however, each blossom contains several stamens and several carpels (or pistil-divisions), and the way in which such a change as this might come about can be easily imagined; for even in many existing plants, where the separate flowers have only a single stamen or a single pistil each, they are nevertheless so closely packed together that they almost form a single compound flower, as in the case of the bur-reed and the various catkins, not to mention the arum and the spurge, where only a trained eye can make out the organic separateness. I shall not trouble you much, however, with these earlier stages in the development of the daisy, both because I shall describe them elsewhere in part, à propos of other subjects, and because the later stages are at once more interesting and more really instructive. It must suffice to say that at some very ancient period the ancestors of the daisy, and of one half the other modern flowers, had acquired an arrangement of stamens and pistils in groups of five,[21] so that each compound flower had as a rule a pistil of five or ten carpels, surrounded by a row of five or ten stamens. And almost all their existing descendants still bear obvious traces of this original arrangement in rows of fives. On the other hand, the ancestors of our lilies, and of the other half of our modern flowers, had about the same period acquired an arrangement in rows of three. And of this other ternary arrangement all their existing descendants still bear similar traces. In fact, most flowers at the present day show clear signs of being derived either from the original five-stamened or the original three-stamened blossom. I don’t mean to say that this is the only mark of distinction between the two great groups: on the contrary, it is only a very minor one;[22] but it is for our present purpose the one of capital importance.
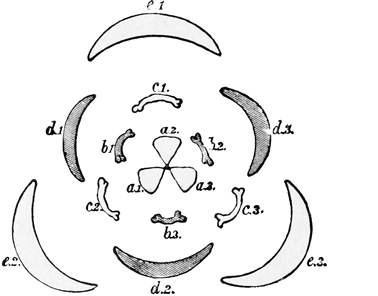
a, Carpels or ovaries; b, stamens, inner row; c, stamens, outer row; d, petals; e, calyx.
Fig. 7.—Diagram of primitive monocotyledonous flower.
The very primitive five-parted common ancestor of the daisy, the rose, the buttercup, and our other quinary flowers, was still an extremely simple and inconspicuous blossom. It had merely green leaves and plain flower-stems, surmounted by a row of five or ten stamens, inclosing five or ten carpels. Perhaps beneath them there may have been a little row of cup-shaped green bracts, the predecessors of the calyx which supports all modern flowers; but of this we cannot be at all sure. At any rate, it had no bright-coloured petals. The origin of these petals is due to the eyes and selective tastes of insects; and we must[23] look aside for a moment at the way in which they have been produced, in order rightly to understand the ancestry of the daisy.
No pistil ever grows into a perfect fruit or sets ripe and good seeds until it is fertilised by a grain of pollen from a stamen of its own kind. In some plants the pollen is simply allowed to fall from the stamens on to the pistil of the same flower; but plants thus self-fertilised are not so strong or so hearty as those which are cross-fertilised by the pollen of another. The first system resembles in its bad effect the habit of ‘breeding in and in’ among animals, or of too close intermarriages among human beings; while the second system produces the same beneficial results as those of cross-breeding, or the introduction of ‘fresh blood’ in the animate world. Hence, any early plants which happened to be so constituted as to allow of easy cross-fertilisation would be certain to secure stronger and better seedlings than their self-fertilised neighbours; and wherever any peculiar form or habit has tended to encourage this mode of setting seeds, the plants have always prospered and thriven exceedingly in the struggle for existence with their less fortunate congeners. A large number of flowers have thus become specially adapted for fertilisation by the wind, as we see in the case of catkins and[24] grasses, where the stamens hang out in long pendulous clusters, and the pollen is easily wafted by the breeze from their waving filaments to the pistils of surrounding flowers. In such cases as these, the stamens are generally very long and mobile, so that the slightest breath shakes them readily; while the sensitive surface of the pistil is branched and feathery, so as readily to catch any stray passing grain of wind-borne pollen.
But there are other flowers which have adopted a different method of getting the pollen conveyed from one blossom to another, and this is upon the heads and legs of honey-eating insects. From the very first, insects must have been fond of visiting flowers for the sake of the pollen, which they used to eat up without performing any service to the plant in return, as they still feloniously do in the case of several wind-fertilised species; and to counteract this bad habit on the part of their unbidden guests, the flowers seem to have developed a little store of honey (which the insects prefer to pollen), and thus to have turned their visitors from plundering enemies into useful allies and friends. For even the early pollen-eaters must often unintentionally have benefited the plant, by carrying pollen on their heads and legs from one flower to another; but when once the plant took to producing[25] honey, the insects largely gave up their habit of plundering the pollen, and went from blossom to blossom in search of the sweet nectar instead. As they did so, they brushed the grains of pollen from the stamens of one blossom against the pistil of the next, and so enabled the flowers to set their seed more economically than before.
Simultaneously with this change from fertilisation by the wind to fertilisation by insects, there came in another improvement in the mechanism of flowers. Probably the primitive blossom consisted only of stamens and pistil, with, at best, a single little scale or leaf as a protection to each. But some of the five-rowed flowers now began to change the five stamens of the outer row into petals; that is to say, to produce broad, bright-coloured, and papery flower-rays in the place of these external stamens. The reason why they did so was to attract the insects by their brilliant hues; or, to put it more correctly, those flowers which happened to display brilliant hues as a matter of fact attracted the insects best, and so got fertilised oftener than their neighbours. This tendency on the part of stamens to grow into petals is always very marked, and by taking advantage of it gardeners are enabled to produce what we call double flowers; that is to say, flowers in which all the stamens have been thus[26] broadened and flattened into ornamental rays. Even amongst wild flowers, the white water-lily shows us every gradation between fertile pollen-bearing true stamens and barren broad-bladed petals. To put it shortly and dogmatically, petals are in every case merely specialised stamens, which have given up their original function of forming pollen, in order to adopt the function of attracting insects.
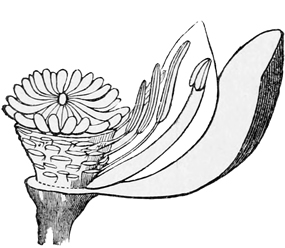
Fig. 8.—Transition from stamen to petal in White Water-lily.
The five-rowed ancestors of the daisy found a decided advantage in thus setting apart one outer row of stamens as coloured advertisements to lure the insects to the honey, while they left the inner rows to do all the real work of pollen-making. They very rapidly spread over the world, and assumed very various forms in various places. But wherever they went, they always preserved more or less trace of their quinary arrangement; and to this day, if you pick almost any flower belonging to the same great[27] division of dicotyledons (the name is quite unimportant), you will find that it has at least some trace of its original arrangement in rows of five. The common stonecrop and its allies keep up the arrangement best of any; for they have each, as a rule, five petals; each petal has its separate bract, making a calyx or flower-cup of five pieces or sepals; inside are one or two rows of five stamens each; and in the centre, a pistil of five carpels. Such complete and original symmetry as this is not now common; but almost all the five-rowed flowers retain the same general character in a somewhat less degree. The buttercup, for example, has one outer row of five sepals, then five petals, and then several crowded rows of stamens and carpels. And in the petals at least the harmony is generally complete. There are five in the dog-rose, in the violet, in the pea-blossom, in the pink, in the geranium, and (speaking generally) in almost every plant that grows in our gardens, our fields, or our woodlands, unless it belongs to the other great division of trinary flowers, with all their organs in groups of three. And now, if you will pull open one of the inner yellow florets of your daisy, you will see that it has five stamens and five little lobes to the bell-shaped corolla, to show its ancestry plainly on its face, and ‘to witness if I lie.’

| Fig. 9.—Corolla of Primrose. | Fig. 10.—Corolla of Harebell. |
But the original bright-coloured ancestor of the daisy must have had five separate petals, like the dog-rose or the apple-blossom at the present day. How then did these petals grow together into a single bell-shaped corolla, as we see them now in the finished daisy? Well, the stages and the reasons are not difficult to guess. As flowers and insects went on developing side by side, certain flowers learnt to adapt themselves better and better to their special insects, while the insects in return learnt to adapt themselves better and better to their special flowers. As bees and butterflies got a longer proboscis with which to dive after honey into the recesses of the blossoms, the blossoms on their part got a deeper tube in which to hide their honey from all but the proper insects. Sometimes this is done, as in the larkspur, the violet, and the garden nasturtium, by putting the honey at the bottom of a long spur or blind sac; and if you bite off the end of the sac in the nasturtium you will find a very appreciable quantity of nectar stored up in it. But most highly specialised flowers have hit upon a simpler plan, which is to run all their petals together at the bottom into a tube, so long that no useless insect can rob the honey without fertilising the plant, and so arranged that the proboscis of the bee or butterfly can rub against the[29] stamens and pistil on the way down. In pinks and their allies we see some rude approach to this mode of growth; for there each petal has a long claw (as it is called), bearing the expanded part at the end; and these claws when firmly pressed together by the calyx practically form a tube in five pieces: but in the perfectly tubular flowers, like the primrose, the arrangement is carried a great deal further; for there we have the claws all grown into a single piece, with the expanded petals forming a continuous fringe of five deeply cleft lobes, representing the five original and separate pieces of the pinks.[2] Now, in the primrose, again, we still find the five petals quite distinct at the[30] edge, though their lower portion has grown together into a regular tube; but in the harebell or the Canterbury bell we see that the whole blossom has become bell-shaped, and that the five originally separate petals are only indicated by five slightly projecting points or lobes which give the tubular corolla its vandyked margin. And if you look at the little central florets of the daisy or the sunflower, you will observe that they too exactly resemble the Canterbury bell in this particular. Hence we can see that their ancestors, after passing through stages more or less analogous to those of the pinks and the primroses, at last reached a completely united and tubular or campanulate form, like that of the heath or the Canterbury bell.
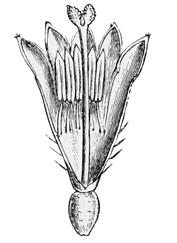
Fig. 11.
Section of floret of Daisy.
There is one minor point, however, in the development of the daisy which I only notice because I am so afraid of that terrible person, the microscopic critic. This very learned and tedious being goes about the world proclaiming to everybody that you don’t know something because you don’t happen to mention it; and for fear of him one is often obliged to trouble one’s readers with petty matters of detail which really make no difference at all except to such Smelfunguses in person. Being themselves accustomed to weary us with the whole flood of their own unspeak[31]able erudition, every time they open their mouths, they imagine that everybody else must be ignorant of anything which he doesn’t expressly state; as though you might never talk of a railway journey without giving at full the theory of kinetic energy as applied to the coal in the furnace. For their sake, then, I must add that, when the daisy’s ancestors had reached a level of development equivalent to that of the heath and the Canterbury bell, they differed in one respect from them just as the primrose still does. In the heath and the harebell, the stamens remain quite separate from the tube formed by the petals; but in the primrose and the daisy the stalks of the stamens (filaments, the technical botanists call them) have coalesced with the petals, so that the pollen seems to hang out in little bags from the walls of the tube itself. This is a further advance in the direction of specialised arrangements for insect-fertilisation; and it shows very simply the sort of cross-connections which we often get among plants or animals. For while the daisy is more like the Canterbury bell in the shape of its corolla, it is more like the primrose in the arrange[32]ment of its stamens. Or, to put it more plainly, while the Canterbury bell has hit upon one mode of adaptation in the form of its tube, and while the primrose has hit upon another mode in the insertion of its stamens, the daisy has hit upon both together, and has combined them in a single flower. And now, my dear Smelfungus, having given way to your prejudices upon this matter, allow me to assure you that nothing will induce me to enter into the further and wholly immaterial difference between hypogynous and epigynous corollas. For every one but you, the very names, I am sure, will be quite sufficient apology for my reticence. These, in fact, are subjects which, like the ‘old familiar Decline and Fall off the Rooshian Empire,’ had better be discussed ‘in the absence of Mrs. Boffin.’
When the ancestors of the daisy had reached the stage of united tubular blossoms, like the harebell, with stamens fastened to the inside wall of the tube, like the primrose, they must, on the whole, have resembled in shape the flowers of the common wild white comfrey, more nearly than any other familiar English plant. The next step was to crowd a lot of these bell-shaped blossoms together into a compact head. If you compare a cowslip with a primrose, you can easily understand how this is done. Accord[33]ing to many of our modern botanists, cowslips and primroses are only slightly divergent varieties of a single species; and in any case they are very closely related to one another. But in the primrose the separate blossoms spring each on a long stalk of its own from near the root; while in the cowslip, the common stem from which they all spring is raised high above the ground, and the minor flower-stalks are much shortened. Thus, instead of a bunch of distinct flowers, you get a loose head of crowded flowers. Increase their number, shorten their stalks a little more, and pack them closely side by side, and you would have a compound or composite flower like the daisy. In fact, we often find in nature almost every intermediate stage: for instance, among the pea tribe we have all but solitary flowers in the peas and beans, long clusters in the laburnum and wistaria, and compact heads in the clovers. The daisies and other composites, it is true, carry this crowding of flowers somewhat further than almost any other plants; but still even here you can trace a gradual progress, some approach to their habit being made by allied families elsewhere; while some composites, on the other hand, have stopped short of the pitch of development attained by most of their race. Thus, certain campanulas have their flowers packed tightly[34] together into a head, which looks at first sight a single blossom, just as deceptively as the daisy does; and a still nearer relative, the scabious, even more strikingly resembles the composite form. So that the daisies and their allies have really only carried out one step further a system of crowding which had been already begun by many other plants.
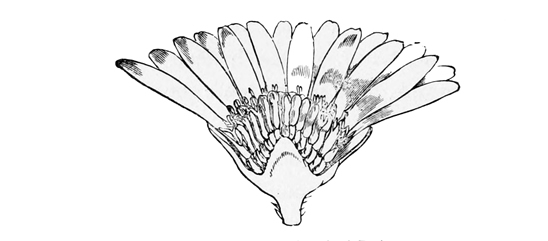
Fig. 12.—Section of head of Daisy.
If you look closely at the daisy, you will see in what this crowding consists. The common flower-stalk is flattened out at the end into a regular disk, and on this disk all the florets are seated with no appreciable separate flower-stalks of their own. Outside them a double row of leaves is arranged, exactly like the calyx in single flowers, and serving the same protective purpose—to preserve the florets from the incursions of unfriendly insects; while inside, the little individual blossoms have almost lost their own calyxes which are scarcely represented by a few tiny[35] protuberances upon the seed-like fruit. In the daisy, indeed, we may say that the true calyx has been dwarfed away to nothing; but in the dandelion and many other composites a new use has been found for it; it has been turned into those light feathery hairs which children call ‘the clock,’ and which aid the dispersion of the seeds by wafting them about before the wind.
Now, what has made the daisy and the other composites grow so small and thick-set? Probably the need for attracting insects. By thus combining their mass of bloom they are enabled to make a great show in the world, and to secure the fertilisation of a great many flowers at once by each insect which visits the head. For each floret has its own little store of honey, its own stamens, and its own pistil containing an embryo fruit; and when a bee lights upon a daisy head, he turns round and round, extracting all he can get from every tiny tube, and so fertilising the whole number of florets at a single time. The result at least proves that the principle is a good one; for few flowers get so universally fertilised, or set their seed so regularly, as the composites. Though they must have reached their present very high state of evolution at a comparatively recent period, they have spread already over the whole world; and they are far more[36] numerous, both in individuals, in species, and in genera, than any other family of flowering plants. In fact, they are undoubtedly the dominant tribe of the whole vegetable kingdom. When I say that in Britain alone they number no less than 120 species, including such common and universal weeds as the daisy, dandelion, thistles, groundsel, camomile, milfoil, hawkweed, and burdock, it will be clear that nine out of every ten ordinary wayside blossoms which we see on any country walk are members of this highly evolved, ubiquitous, and extremely successful family.
Still, we are far from having finished the pedigree of the daisy. We have traced its general genealogy down as far as the common composite stock: we have now to trace its special derivation from the early common composite type to the distinctive daisy form. Clearly one great point in the daisy’s history is yet untouched upon; and that is the nature and meaning of the white rays. We know that the inner yellow florets are (as it were) dwarfed and specialised golden harebells; but we do not yet know what is the origin of these long outer streamers, which look so wholly unlike the tiny and regular central bells.
In solving this problem, the other composites will help us not a little; for we must always seek in the simpler for the interpretation of the more complex;[37] and the daisy, instead of being the simplest, is one of the most developed representatives of the composite pattern. If you turn to that tall, rank-looking weed growing yonder, under cover of the hedge, you will get a good surviving example of the earliest form of composite. The weed is a eupatory—‘hemp agrimony’ the country people call it—and it has small heads, each containing a few tubular purple florets, all exactly the same size and shape, and all much more loosely gathered together than in the daisy or the dandelion. The eupatory is interesting as preserving for us one of the first stages in the ancestry of the higher composites, after they had attained to their distinctive family characteristics. Once more, I don’t wish you to understand that the daisies are descended from the eupatory: all I mean is, that their ancestors must once have passed through an analogous stage; and that the eupatory has never got beyond it, while the daisies have gone on still further differentiating and adapting themselves till they reached their present peculiar form. Now, if you compare this daisy with the head of eupatory, you will see that they differ in two particulars—the daisy has outer rays, while the eupatory has none; and the inner daisy florets are yellow, while the eupatory florets are purple. The latter difference is[38] one into which we cannot enter now: it must suffice to say that when the daisy’s ancestors were in the eupatory stage of development they had apparently all their florets yellow. This is likely, because almost all the modern composites of every sort have yellow central florets, and most of them have yellow rays as well. It is only a few kinds that have red or purple central florets; and, as we shall soon see, only a few also that have white or pink outer rays.
What, then, made the daisy’s ancestors produce a row of external florets so different in shape and colour from the internal ones? The answer is exactly analogous to that which I have already given for the origin of petals themselves. Compare the eupatory with the daisy once more, and you will see that the one is comparatively inconspicuous, while the other is very noticeable and bright-coloured. The row of green bracts almost hides the blossoms of the eupatory; but the large white rays make a bold and effective advertisement for the daisy. Certain composites, in fact, have just repeated the same device by which the earliest petal-bearing flowers sought to attract the notice of insects. Those early flowers, as we saw, set apart one outer row of stamens as bright-coloured petals; these later compound flower-heads have set apart one outer row of florets as bright[39]-coloured rays. If you examine the rays closely, you will see that each of them is a separate little flower, with the stamens suppressed, and with the bell-shaped corolla flattened out into a long and narrow ribbon. Even these very abnormal corollas, however, still retain a last trace of the five original distinct petals; for their edge is slightly notched with five extremely minute lobes, often nearly obliterated, but sometimes quite marked, and almost always more or less noticeable on a careful examination. A daisy thus consists of a whole head of tiny tubular bells, the inner ones normal and regular, with corolla, stamens, and pistil, and the outer ones flattened or ligulate, with the stamens wanting, and the entire floret simply devoted to increasing the attractiveness of the compound mass. Pull off the rays, and you will see at once what an inconspicuous flower the daisy would be without them.
Last of all, the question arises, Why are the outer florets or rays pink and white, while the inner florets or bells are golden yellow? When we have solved that solitary remaining problem, we shall have settled the chief points in the daisy’s pedigree. Clearly, when the rays were first produced, they must have been yellow like the central florets. The mere flattening and lengthening of the corolla would not[40] in itself tend to alter the colour. And as a matter of fact, the vast mass of those composites which have progressed to the stage of having rays—which have got these two separate forms of flowers, for show and for use respectively—have the rays of the same colour as the central bells, that is to say, generally yellow. Of this stage the sunflower is a familiar and very striking representative. It has bright golden central florets, and large expanded rays of the same colour. To anybody who wants to study the structure of the daisy without a microscope, the sunflower is quite as valuable and indispensable as it is to our most advanced æsthetic school in painting and decoration. Moreover, it shows us admirably this intermediate stage, when the compound flower-head has acquired a distinct row of outer attractive florets, adding wealth and expansiveness to its display of colour, but when it has not yet attempted any specialisation of hue in these purely ornamental organs. The daisy, however, together with the camomile, the ox-eye daisy, and many other similar composites, has carried the process one step further. It has coloured its rays white, and has even begun to tinge them with pink. This makes these highest of all composites the most successful plants in the whole world. If one considers that daisies begin to bloom on January 1, and[41] go on flowering till December 31; that they occur in almost every field far more abundantly than any other blossom; and that each one of them is not a single flower, but a whole head of flowers—it will be quite clear that they are much more numerous than any rival species. And when we add to them the other very common white-rayed composites, such as the camomiles, many of which abound almost as freely in their own haunts and at their proper season, it is obvious that this highly evolved composite type is the dominant plant race of the old world at least. In the new world, their place is taken by a somewhat more developed type still, that of the Michaelmas daisies, which have their rays even more ornamental than our own, and brightly coloured with mauve or lilac pigment. All the world over, however, in and out of the tropics, the commonest, most numerous, and most successful of plants are ray-bearing composites of one kind or another, like the daisies, with the rays differing in colour from the central florets.
Finally, it may, perhaps, at first hearing, sound absurd to say that the daisy group, including these other composites with tinted rays, forms the very head and crown of the vegetable creation, as man does in the animal creation: and yet it is none the less true. We are so accustomed to look upon a daisy as a[42] humble, commonplace, almost insignificant little flower, that it seems queer to hear it described as a higher type of plant life than the tall pine-tree or the spreading oak. But, as a matter of fact, the pine is a very low type indeed, as is also the giant tree of California, both of them belonging to the earliest and simplest surviving family of flowering plants, the conifers, which are no better, comparatively speaking, among plants, than the monstrous saurians and fish-like reptiles of the secondary age were among animals. If size were any criterion of relative development, then the whale would take precedence of all other mammals, and man would rank somewhere below the gorilla and the grizzly bear. But if we take complexity and perfection in the adaptation of the organism to its surroundings as our gauge of comparative evolution, then the daisies must rank in the very first line of plant economy. For if we follow down their pedigree in the inverse order, we shall see that, inasmuch as they have coloured rays, they are superior to all their yellow-rayed allies (for example, the sunflower); and inasmuch as these have rays, they are superior to all rayless composites (for example, the eupatory); and inasmuch as composites generally have clustered heads, they are superior to all other flowers with separate tubular corollas (for example,[43] the heathers); while all these, again, are superior to those with separate petals (for example, the roses); and all petalled flowers are superior to all petalless kinds (for example, the pines and oaks). Thus, from the strict biological point of view, it becomes quite clear that the daisies, asters, chrysanthemums, and other rayed composites with coloured outer florets, really stand to other plants in the same relation as man stands towards other animals. That is what gives such a special and exceptional interest to the daisy’s pedigree.

Fig. 13.—Hairy Wood-spurge (Euphorbia pilosa).
You will not find many pleasanter or breezier walks in England than this open stretch of Claverton Down: certainly you will find very few with more varied interest of every conceivable sort for every cultivated mind. The air is fresh and laden from the brine of the Atlantic and the Gulf Stream; the clear wind is blowing straight from seaward, not keen and dry from the Eastern plains, but soft and pure from a thousand leagues of uninterrupted ocean; and the view over the broken dale of Avon, where it cuts its way in a veritable gorge through the high barrier of the Bath oolite, stretches for miles over one of the loveliest and greenest valleys in all our lovely green England. More than that—the whole history of Britain is visibly unfolded before my very eyes. That bald roundish hill to the right, with its smooth summit artificially levelled, and its sides planed down into a long glacis, is Little Solisbury; and Little Solisbury, as its name clearly shows, is the very oldest Bath of all. For it is[45] the bury or hill-fort of Solis, the ancient fortified town of the Keltic and Euskarian natives; and when, long ages afterwards, the Romans planted their station in the valley below, they naturally called the hot springs which they found there by the name of Aquæ Solis; and equally naturally misinterpreted the second word (really a native term, Sulis) as the genitive of Sol, and accordingly dedicated their great temple on the spot[46] to Apollo. Those straight white lines and green-grown ridges on the flanks of Banagh Down and the eastern heights are the vestiges of the old Roman causeways—the Fosse and its branches—now totally disused or else degraded into modern cart-roads; and the Institution Buildings in the valley below cover or contain all the remaining memorials of the stately Roman town. Back of me again, on Hampton Down, stand the earthworks of Caer Badon, the later British village, planted there when fear of the heathen West Saxon invaders had driven back the Christian Welshman to the hills which he had deserted for the fruitful valley during the security of the Pax Romana; and this long mound, on whose summit I am standing to catch the view, actually forms part of Wansdyke, the great boundary barrier behind which the Welshmen of the Somersetshire principality entrenched themselves, after the pagan English pirates had taken possession of the Avon dale and of Bath itself. The decisive battle which settled the fate of the city was fought at Dyrham Park, among those blue downs on the northern horizon; and the tiny village of Englishcombe, nestling below the solitary beacon of High Barrow Hill on my left, marks in its very name the furthest westward extension of the Teutonic settlers towards the ever-unconquered recesses of Mendip.[47] As to later associations, they are too endless for review. In the foreground lies the town, and from its midst towers the abbey, the last flickering effort of English architecture before the Reformation choked out its life for ever; a tall and stately but very cold specimen of good late perpendicular work. It rises above the ancient temple of Minerva, and covers fragments of the older minsters—that which Osric, king of the Worcester men, gave to a nunnery in 671; that which Offa of Mercia raised in 775; that where Eadgar, first king of all England, was crowned in 973: and that which the Angevin John of Tours erected in 1160. There to the right is Lansdown, where the Parliament’s men under Waller all but wiped out the stout Cornishmen who ‘stood up for their king’ under Sir Bevil Grenville in a fruitless victory; and the big tower on the top is Beckford’s Folly, built in a fit of Oriental recklessness by ‘Vathek’ Beckford, and now the landmark of the cemetery which spreads over his vanished domain. In the combe to the left, again, that huge pseudo-classical manor-house is Prior Park, the vast rambling home of Ralph Allen; and Ralph Allen was the original of Squire Allworthy, whose grounds, as minutely described in ‘Tom Jones,’ are here actually realised. But if I went on talking all day I should never have[48] finished; for the history of the Bath valley, as seen from Claverton Down, is, as I said before, the history of all England, visibly epitomised in tangible realities before one’s very eyes.
However, I have not come out to-day to hunt for old relics among the works of Caer Badon, or to trace the curious bends and angles of Wansdyke. A far older and stranger chapter of our history than any of these is unfolded by the little wayside weed which I have here in my botanical case; and it was to find this very commonplace and uninteresting-looking plant that I have come out this morning. For the weed is the hairy wood-spurge, and Claverton Down is the only place in Great Britain where that particular kind of spurge still lingers on. I have got my British Flora safe here in my satchel; and now I am going to sit down on the slope of Wansdyke and make quite sure that my plant really tallies exactly with Dr. Bentham’s description; for if it actually does, then I shall have the pleasure of knowing that I hold in my hand one of the few genuine links which yet unite us with a very distant past—a past compared with which the days when Wansdyke was built, or even when Little Solisbury was fortified, seem comparatively recent. If this is in fact the hairy wood-spurge,[3] it and its ancestors have been growing here[49] on Claverton Down ever since the end of the last glacial epoch; and it is a relic of the flora which once bloomed among the lowlands that connected England and Ireland with Brittany, Spain, and the Pyrenees. It dates back, in short, to the time when Britain was still an integral part of the European continent.
A few minutes’ examination with my pocket-lens is quite enough to assure me that the flower I have picked is truly the wood-spurge of which I am in search. It is a queer, insignificant little plant, with funny cup-like green flowers, and odd jelly-bag glands, very much like most other English spurges; but I see at once on a closer examination that it has all the distinguishing marks of the hairy species—the woolly underside to the leaves, the dotted seed-capsules, the loose umbels of blossom, and the long branched rays supporting the straggling flower-heads. I regard it, therefore, as a decided find; for the lane that bounds the Prior Park estate, and this bit of woodland on the summit of Claverton Down, are the only spots in England where this particular plant is now found. But that is not all. In itself, the fact of its rarity would not be enough to arouse any special interest; for there are many other wild flowers found in only one spot in Britain—sometimes garden kinds[50] escaped from cultivation in a suitable climate, sometimes American straylings, and sometimes high Alpine species requiring a particular granite, basalt, or limestone soil—a soil perhaps to be met with in our islands only on one or two scattered Welsh or Scottish hills of the requisite height. The case of the hairy spurge, however, is very different from any of these. It is a southern European and Western Asiatic plant, and it spreads along the Mediterranean basin from the Caucasus to the Pyrenees; but it nowhere comes any nearer to Britain than the valley of the Loire. This is what gives it such a special interest in my eyes. It is not found in Brittany, it is not found in Normandy, it is not found on the opposite coast of Picardy, it is not found in Kent or Essex; but it suddenly reappears here, out of all reckoning, on Claverton Down.
If the case of the wood-spurge were a solitary one, it would be easy enough to give a ready explanation. The neighbourhood of Bath is known to be one of the warmest spots in England, having, in fact, its own hot-water supply always laid on. This is a plant of warm countries. A bird, let us say, once brought over a single seed, clinging to its feet or feathers; an exotic flower, imported for the shrubberies of Prior Park, was packed in earth containing young spurges;[51] a sailor introduced it by some chance; a botanist sowed it here for an experiment. Nay, perhaps a Roman settler at Aquæ Solis brought it over with the plants for his Italian garden. In such or the like casual manner it got a footing on Claverton Down; and, as the climate suited it, it has gone on flourishing ever since. Here, I say, would be an easy explanation if the case of the hairy spurge were a solitary one; but, as a matter of fact, there are hundreds of cases exactly like it. It is quite a common occurrence to find a plant extend all through Europe from the Caucasus to the Pyrenees, then stop suddenly short, and turn up again once more incontinently in Devon, Cornwall, Kerry, and Connemara. This is such a curious fact that it really seems to call for some adequate explanation.

Fig. 14.—Flowers of common Monkshood.
Let me begin by noting a few of the most striking instances. There is in the Bristol Channel a solitary rocky islet known by the old Scandinavian title of the Steep Holme—a name given to it, no doubt, by the wickings of the ninth century, who made it their headquarters for plundering the chapmen and slave-mongers of wealthy Brycgstow. Now the rocky clefts of the Steep Holme are still crimson in May and June with the brilliant red blossoms of the wild pæony, a flower which does not elsewhere appear[52] nearer to England than the Pyrenees. Not far from Axminster in Devon, again, there is a warm sheltered nook in which nestles the little village of Kilmington. Well, Kilmington Common is a place famous to botanists, because it is the one single station in Britain for a small purplish lobelia, which ranges elsewhere only from Andalusia to central France.[53] Dozens of like cases may be noted in the south-western peninsula of England and the similarly situated corner of Wales about Pembrokeshire. Thus, to lump a long list briefly, the common blue monkshood is found wild in South Wales and the Cornish district only; the yellow draba is confined to old walls about Pennard Castle, near Swansea; the spotted rock-cistus occurs only in the Channel Islands and at Holyhead; the white rock-cistus is peculiar in Britain to Brent Downs in Somerset, together with Torquay and Babbicombe in Devon; the Cheddar pink, a volcanic plant of southern Europe, clings to the crannies of the Cheddar cliffs near Wells, and to no other crag in England; the soapwort is wild only in Cornwall and Devon; the flax-leaved St. John’s wort grows nowhere but at Cape Cornwall and on the banks of the Teign; the crimson clover and Boccone’s clover are entirely restricted to the peninsula of the Lizard; so also is the upright clover, save that it is likewise found in the Channel Islands; the sand bird’s foot remains only at Scilly; the Bithynian vetch extends through Europe as far north as Bordeaux, and then disappears again till after a sudden leap it is gathered once more in Devon and Cornwall; the white sedum occurs in the Malvern Hills and in Somersetshire; and the narrow buplever flowers only[54] at Torquay and in Jersey and Guernsey. In almost all, if not in all, these cases the plant is a southern one, which extends usually from the Caspian to Spain, is perhaps found as far north as the Gironde or even the Loire, and then disappears again till it turns up suddenly in some exceptionally sheltered nook of Devon, Cornwall, or South Wales. This is a phenomenon which cannot surely be due to chance alone. Indeed, I might greatly increase the list, but I refrain only because I am afraid of being wearisome.
When we turn to the similarly placed south-western corner of Ireland, the peculiarities we meet are even more remarkable. I shall never forget my surprise when once, after my first visit to Nice and Mentone, I began describing the beautiful Provençal flowers to an Irish botanist, and was quietly answered, ‘Ah, yes; we have them all at Killarney.’ But it is really true none the less. The thick-leaved sedum, after skipping all England and Wales, shows itself suddenly in the Cove of Cork. The pretty Mediterranean heath, which every winterer at Pau has gathered by handfuls on the hills about Eaux Chaudes or Cauterets, jumps at a bound to the coast of Kerry. The arbutus, with its clustering white blossoms and beautiful red berries, is similarly found in Provence and again at Glengariff. London Pride[55] grows wild in Portugal, western Spain, and the higher Pyrenees, and reappears in south-western Ireland. Another pretty little saxifrage jumps in like manner from the Asturias to Killarney. St. Dabeoc’s heath has the same range. The spiked orchid takes a great bound from Bordeaux to a single station in County Galway. To sum it up shortly, ‘Crete, Auvergne, the Pyrenees, S.-W. Ireland,’ is a common technical description of the distribution of many beautiful south European plants.
Fig. 15.—Flower and fruit of Arbutus.
Now, these peculiarities of distribution lead me up pretty surely to the romance of the hairy wood-spurge. They show that it did not get here by accident. Like the elephant-headed god of the Mexicans, like the debased traces of Buddhism in the Aztec religion, they raise an immediate curiosity as to their origin. What we may call the natural range of British plants is of this sort: they have entered the country from the Continent, viâ Kent, Sussex, East Anglia, or Scotland; and they fall for the most part under three great divisions. The first division consists of central Euro[56]pean plants, which seem as if they had come in from the east: and of these a few get no farther than the eastern counties; a great many spread over the whole country; and the remainder have reached to the west and to Ireland. The second division is that of the Scandinavian plants, which seem as if they had come in from the north; and of these a few stop short in Shetland, Orkney, or the Highlands; others get as far as the midland counties; and a good many straggle on into Kent or Cornwall. The third division comprises the mountain plants, which have come in from various quarters, and which grow wherever the elevation and the mountain air suit their constitutions. But my wood-spurge agrees with none of these, and it clearly belongs to another southern class, which cannot have entered Britain by any of the customary routes viâ Dover, Harwich, or Southampton. It seems to have taken a route of its own, and to have attacked England by way of Bristol and Bordeaux. Otherwise, we should find it and the other peculiar west-country species in the warmer parts of Kent, Surrey, and the Isle of Wight, which, as a matter of fact, we never do. If climate were the only agent at work, Ventnor certainly has as good claims as any place in England.
Perhaps it seems a useless question to inquire how[57] they came there at all. ‘Were they not always there?’ somebody may ask me. And the answer is, No, undoubtedly not. You might as well explain the presence of an English-speaking colony on Pitcairn Island by the hypothesis that Englishmen were originally created in two separate centres—Great Britain and the South Pacific. Only some 80,000 years since—a mere single swing of the cosmical pendulum—every inch of Great Britain and Ireland, save only an insignificant southern fringe, was wholly covered by the ice of the last glacial period. We know the date with mathematical certainty, because the astronomical conditions upon which glacial periods have been shown almost beyond doubt to depend, began 200,000 years ago, and ended 80,000 years ago. During the interval between those two dates, the condition of each hemisphere alternated between long cold periods and long hot periods, of some 10,500 years each. During the last cold spell, all England and Ireland were in the condition of Greenland at the present day. The ice had planed every living thing clean off the face of the country, and we may still trace its scratches on the smooth granite bosses of Wales and Scotland, or find its till and its moraines on the plains and valleys of East Anglia and Derbyshire. Consequently the ancestors of every[58] plant and every animal now living in Britain must have come into it after the end of the last long cold spell—that is to say, roughly speaking, some 80,000 years since.
Moreover, when Britain was repeopled after the great ice age, it must have been united to the Continent somewhere, or else it could not possibly possess the large number of European plants and animals which it actually contains.[4] Had it then been an island, it might have had a considerable population of ferns and small-seeded flowers, of birds and winged insects, blown over to it from the shores of France or Holland; it might even have had a fair sprinkling of snails and lizards, or a few small quadrupeds, wafted across on logs of wood, or carried over accidentally by various chances; but it would be quite impossible that it should have all the species of large or middle-sized wild mammals which we see now inhabiting it—the red deer, the fallow deer, the otter, the badger, the fox, the hare, the rabbit, the weasel, the stoat, the marten, the hedgehog, the wild cat, the mole, the shrew, the squirrel, and the water-vole. Altogether, we have no less than forty species of British mammals; while the bear, the wild boar, the beaver, the reindeer,[59] and the wolf have become extinct within the historical period; and the wild white cattle even now survive sparingly in Chillingham Park and a few other scattered places. Clearly, as none of these animals or their ancestors can have been in Britain 80,000 years ago, they must have come into Britain at some later date, across a wide bridge of solid land. For Mr. Wallace has conclusively shown that islands which have never formed part of a mainland never have any terrestrial mammals at all; and that a very narrow strait is quite sufficient to prevent the passage of mammals from one island to another. The sound which divides the Indo-Malayan region from the Australian region is hardly wider than that which separates England from France; yet not one single Australian mammal has ever reached the Indo-Malayan region, and not one single Indian mammal has ever reached Australia. The kangaroos, wombats, phalangers, and cassowaries of the one district are quite distinct in type from the elephants, tapirs, tigers, deer, and monkeys of the other. So that our numerous existing English fauna must certainly have crossed over on dry land.
We may take it for granted, then, that the mass of British plants came in, from the east and south-east, immediately after the ice of the glacial epoch had[60] passed away. For the ice had driven man and beast, herb and tree, southward before it; and even if there was a little fringe of what is now Southern Britain not wholly glaciated, yet its condition must have been like that of the little habitable fringe in Greenland, and its plants and animals (if any) must have been of thoroughly Arctic types. But as the glaciers cleared away again, with the return of the sun to the northern hemisphere after its long cold cycle, the southern and eastern plants and animals must have followed the retreating ice-sheet from year to year; till at last the species which used to inhabit Kent and the Isle of Wight found their permanent home in Lapland, and those which used to inhabit Greece and Italy found their permanent home in Holland, Denmark, and Great Britain.
This sufficiently accounts for the presence in England and Scotland of the central European and Scandinavian elements; but it does not account for the presence of my hairy spurge and of all the other south-western species, belonging to the Pyrenean and Italian region. Clearly, the ordinary plants of Eastern England are plants which once spread uninterruptedly from Warwickshire to Central Europe, when the belt of land over the German Ocean was still entire; and clearly, too, the ordinary plants of the North and of[61] Scotland are plants which once spread uninterruptedly from Yorkshire to Scandinavia, during the same period; while both classes have been afterwards isolated in Britain by the gradual subsidence of the intervening land. But this still leaves unanswered the question, Whence did we get the Pyrenean types?
Perhaps one might be disposed at first sight to fancy that they came over separately, as we know a few American plants have really done. There is the well-known Canadian canal weed, which was introduced by a botanist into a tank near Cambridge in 1845, and rapidly spread over all England; there are a few orchids and other wild flowers whose seeds have apparently been carried across the Atlantic on the feet of birds; and there are some half-dozen escaped garden flowers, like the evening primrose, which have established themselves easily among some rare warm spots in our congenial climate. Possibly it might seem as though the arbutus, the hairy spurge, the Mediterranean heath, and all the rest of the southern species in South-Western England or Ireland had got across to us in somewhat the same fragmentary fashion, and had succeeded in effecting a foothold only in these warmer Cornish and Irish nooks. But there are a great many reasons against believing this. In the first place, we have the immense number to[62] account for—at least ninety species, all told; which is a prodigious item to set down to the chapter of accidents. For the distance from Bordeaux to Kerry is really 700 miles, while the distance from Portugal to the Azores (which are peopled with plants and animals in the most fragmentary manner) is only 900; and we can hardly suppose that so large a number of southern plants could permanently establish themselves (against the prevailing winds) in a country already occupied by a flourishing native flora. But two more fatal objections are these: First, our southern plants are only found in the extreme south-west, and not in the warmest parts of the Isle of Wight, of Kent, or of Hampshire. Even at Bournemouth and Ventnor we meet with none of them. And secondly, they are all evidently dying out; they represent an old flora which is no longer adapted to the country, not a new flora pushing its way vigorously into regions occupied by less congenial plants. Every year they are disappearing before our very eyes, and many of them are from time to time now being expunged from our floras. The Kilmington lobelia is getting rarer as every summer passes; the wild asparagus, once common on the Lizard promontory, is now only to be picked, at the imminent risk of life and limb, amongst the crannies of a rocky islet at Kynance Cove; the[63] purple viper’s-bugloss has been driven to the very extremity of Britain at Penzance; while the various kinds of rock-cistus, the Steep Holme pæony, and the Cheddar pink linger on each only in a single inaccessible spot in the south-western peninsula of England. These are clear evidences that they form the last stragglers of a vanquished flora, not the vigorous vanguard of a victorious and aggressive race.
And now we are in a position fairly to settle the problem where the hairy spurge and its fellows have come from, and how they got here. People who recognise the fact that Britain was once joined to the Continent are too apt to fancy that it was joined only by a sort of narrow bridge between Dover and Calais. The aspect of the shore on either side, the high bluffs of Shakespeare’s Cliff and Cap Grisnez, the geological continuity between the chalk and the other formations on the two coasts, all forcibly suggest that France and England must once have been joined there—as, indeed, they undoubtedly were. But we are all inclined mentally to minimise the amount of connection; we stick in an isthmus just sufficient to carry the South-Eastern Railway across to Boulogne, and then we are fully satisfied with our new geography. In reality, however, the old land connection was something far more complete and universal than that.[64] There is every reason to believe that, at the close of the last glacial epoch, Great Britain and Ireland formed a part of the Continent, not in the sense in which Scandinavia or Denmark still does, but in the sense in which Bavaria and Switzerland still do. The land of Europe then stretched out to seaward far beyond Ireland, Spain, and the Faröe Islands; and Cork, Glasgow, and Liverpool then stood further inland than Lyons, Munich, and Geneva stand at the present day.
Walking one morning a few winters since—just after the most terrible tempest of recent years—on the Parade at Hastings, I happened to notice a curiously shaped flint among the shingle lately thrown up by the great storm. The waves had beaten right over the sea-wall, and scattered a litter of wrack and pebbles along the whole roadway. I stooped down and picked up the odd-looking fragment: to my surprise, I found it was a palæolithic implement, a rudely chipped flint knife of the older stone age, the relic of a race compared with whom even the builders of Wansdyke here were men of yesterday. This rude flake was fashioned by the naked black-fellows who hunted the rhinoceros and the mammoth in the English valleys, before ever the great ice age itself had spread its glaciers over the length and breadth of[65] the land, a couple of hundred thousand years since. Its outer surface was dulled and whitened by age, as is always the case with these primæval flint weapons; but its edge was still sharp and keen, though crusted in places with a hard film of mineral deposit, and also blunted here and there by use in cutting clubs and reindeer bones for its savage possessor. But there were no traces of rolling as in water-worn pebbles: the knife was freshly disinterred. It was clear that the storm had just unearthed it from beneath the submerged forest which belts all the coast from Beachy Head to Dungeness. For the forest is a post-glacial deposit; and it once formed part of this great connecting land, now buried beneath the Atlantic, the English Channel, and the German Ocean. The trees which composed it still stand as upright stumps, firmly bedded in a layer of tenacious clay; and strewn beneath them lie prostrate boles, in the very place where the wind threw them down some fifty or sixty thousand years ago. In the public garden at Hastings, one of these huge balks, dug up on the St. Leonard’s beach, has been fixed as a curiosity; and, though its outer layer is charred and blackened by the water, the inner wood is still as sound and as firm as on the day it fell. We have to deal here with a time which is marvellously ancient indeed when measured[66] by our ordinary human and historical chronology, but which is quite modern when judged by the vast timepiece of cosmical and geological cycles.
All round the coast of England you will find endless traces of these submerged forests, especially wherever the land shelves off slowly to seaward. That most lively of mediæval travellers, Giraldus Cambrensis (whose amusing and somewhat slangy diary would be much more read, I am sure, if people did not incongruously mistake him for a dry chronicler of the monastic sort), gives a full and really scientific account of one which he came across in the course of his Welsh peregrinations; and ever since his time the submerged forests have been noted in spot after spot in every part of Southern Britain. Beginning in the great bight between Wales and Scotland, they continue round the coast at Holyhead; turn up again in Cardigan Bay; fringe the whole Bristol Channel; fill in the bottom of the fiords at Falmouth, Dartmouth, Torquay, and Exmouth; trend round the Isle of Wight, Selsea, and Pevensey Bay; appear sparingly off the Essex coast; and thence run up by Cromer and the Wash to Holderness and Lindisfarne. They are everywhere newer than the glacial deposits, and so they give us a fair ground for believing that a great general subsidence of the land has taken place[67] all round the shore of England at a comparatively recent period—that is to say, since the close of the last glacial epoch. How recent they are is well shown by the nature of the remains themselves; for they often contain undecayed leaves, water-logged hazelnuts, bits of small twigs, and other forestine rubbish of a perfectly undecayed and modern-looking character. Some of the twigs even break with a sharp crackling sound, like dry wood freshly taken from a modern forest.
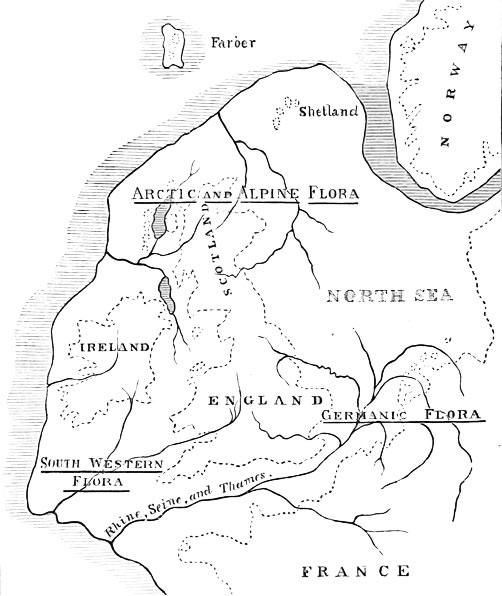
Fig. 16.—Sketch Map of Post-Glacial Britain.
The question now remains, If the land once thus extended farther out to sea than at present, how far out did it extend? or, in other words, how large a subsidence has taken place? Here we have an excellent hint for our guidance in the fact that Ireland must have been united to England since the glacial epoch, because we find in Ireland a large proportion of the British plants and animals, including a considerable number of land mammals. Now, how much must we raise the general land surface of the British Isles in order to unite Ireland to Great Britain? Well, a rise of less than one hundred fathoms would suffice to join the whole of our islands throughout nearly all their length, leaving only two large lakes in the very deepest parts of the sea, where the plummet marks a depth of a hundred and fifty fathoms. One[68] of these two large lakes would lie between Galloway and Ulster, and the other would fill up the hollow of the Minch between the Hebrides and the Isle of Skye. But the same amount of elevation would also suffice to unite us to the Continent from Denmark to Spain, as well as to push out our whole coast-line about fifty[69] miles to the westward of Cape Clear. Beyond that distance the sea-bottom suddenly topples over from a general depth of a hundred fathoms to a depth of a thousand fathoms or more; which clearly shows that this line, curving round from Shetland to the Spanish shore of the Asturias, must mark an old and long-continued prehistoric land-barrier. In other words, the British Isles are situated on a comparatively shallow submarine bank, which spreads north, south, and east of them, but ends abruptly to the westward by a sudden drop of eight or nine hundred fathoms. If you were now to raise this bank a hundred fathoms in height, you would lift its whole area above the sea-level, save only in the two hollows already noted; but if you went on raising it for several hundred fathoms more, you would not materially alter the coast-line established by your first elevation. So we can hardly doubt that the hundred-fathom line really represents the old western boundary of Europe towards the Atlantic, because it coincides so nearly in depth with the elevation necessary to unite England and Ireland to one another, and to the Continent.
Only one element of our problem now remains to be solved; and that is the question—When did the subsidence take place which turned the dry land all round Britain into the beds of the English Channel,[70] the German Ocean, and the Irish Sea? To this question I am deferentially inclined to give a somewhat different answer from that of most of our authorities. As a rule, it seems to be implied that the subsidence was a single act, spread indeed over a considerable length of time, but completed once for all, and never since renewed. It appears to me more probable, however, that the subsidence has been going on more or less ever since the age of the submerged forests, and that it still continues in places over the same area. Mr. Wallace has already pointed out that Ireland was probably separated from the mainland sooner than England, because it has fewer native mammals and hardly any reptiles or amphibians. The happy immunity from toads and serpents which is generally attributed to the pious exertions of St. Patrick, may perhaps rather be set down to the early isolation of Ireland from the mainland shortly after the end of the great ice age, and before all the members of the new European fauna had had time to spread equally over the more outlying portions of the yet undivided continent. But there are other indications of subsequent partial submergence elsewhere. Many facts lead me to the belief that the Bristol Channel was still a plain through which the Severn flowed quietly to the sea long after the final insulation[71] of Ireland and the Hebrides. Tourists driving from Barmouth to Port Madoc have looked down from the picturesque escarpment of Harlech Castle upon a narrow belt of plain between the mountains and the sea, and have been told how the Lowland Hundred once stretched outward from this point across Cardigan Bay as far as Sarn Badrig or St. Patrick’s Causeway, a rocky reef which whitens the Channel into a long line of breakers in the middle distance. Welsh legends, immortalised by Peacock’s delicious satire, tell us how the Hundred was submerged by an inundation; and the tradition as to this subsidence is almost certainly correct. There is some ground for believing that the Isle of Wight was still united at ebb tide to the mainland of Hampshire by a sandy isthmus, when the Romans built their villas at Brading; and we know that even as late as the days when Hengest and Horsa launched their mythical long ships for the conquest of Kent, the Zuyder Zee was yet undoubtedly an inland lake, separated from the German Ocean by a long belt of land now almost entirely submerged, save in the solitary line of islands which preserves the outline of its northern shore. Nay, even in our own time, the southern part of Sweden is slowly sinking by inches beneath the level of the Baltic. Hence I am strongly inclined to sus[72]pect that the submergence of this western land was a work of time, and that no particular date can be assigned to it as a whole.
Now, when a continuous belt of lowland stretched round from Spain to Ireland and the Shetlands, we can easily understand that the warm type of south European plants would run northward along its western shore as far as the climatic conditions permitted. But the climate on all the west coast of northern Europe is exceptionally mild and moist, through the agency of the Gulf Stream and the warm westerly breezes which blow across it. Hence it is not surprising that the Mediterranean heath, the strawberry tree, the pæony, the hairy spurge, and all the other southern plants which I have before scheduled, should have ranged all along the Atlantic shore of Europe, past the Pyrenees and the Asturias, up the bend a hundred and fifty miles west of the Land’s End, and so onward to Kerry and Connemara. Dr. James Geikie has recently shown good reasons for believing that the last glacial epoch was immediately succeeded by a short spell—say a thousand years or so—of very sunny and genial conditions in northern Europe; and while these favourable conditions lasted we can readily understand that the southern flora may even have extended along the[73] sheltered belt beneath the mountain-ranges of Ireland and Scotland as far northward as Bute and Arran, where some few of its hardier representatives are actually still preserved. Meanwhile, the eastern level slope of what is now England, together with Holland and the intervening land which then filled up the basin of the German Ocean, must have had an inland continental climate, exposed to the full rigour of the north-east winds, and unmitigated by the warmth and moisture now diffused over it by the sea and its currents. In short, the condition of that great tableland must have been much like the condition of Central Russia at the present day, aggravated perhaps by an extra elevation to some hundreds of feet above its existing level. Here, then, the flora must have been of the central European and Scandinavian type; while west of the great central range of England, the trees and flowers must in the main have resembled those which we now find among the nooks of the Apennines and the Genoese Riviera.
By-and-by, however, the earth’s crust began to sink in western Europe, as it is sinking now in Scania and the bed of the southern Baltic. Slowly the great Atlantic plain disappeared below the waters, leaving only the mountain-tops and higher plateaus as islands above the sea-level. First the two lateral[74] valleys of the old lake-system were flooded, cutting off Ireland and the western Hebrides as two large and compact islands, considerably bigger than they now remain at the present day. Then, doubtless, the North Sea and the Channel were overflowed, leaving only a narrow neck of chalk downs as a connecting link between Kent and Picardy, which the waves gradually beat down and at last destroyed. The cliffs of Dover and Cap Blancnez, of Beachy Head and Dieppe, now mark its limits. Still the Bristol Channel remained an open valley, and Scilly was united to the Cornish peninsula. Next, Scilly and the Channel Islands went; while the Hebrides and the western coast of Scotland broke up into a number of separate islets, only the granite crests of the higher mountain-ranges now overtopping the water in long lines, while the lateral valleys became the straits which separate the various members of the different archipelagos from their nearest neighbours. Any one who has once yachted down the broken ridge of the Outer Hebrides cannot fail to have noticed that they seem but the summits of a vast sunken range, jagged and beaten at the outer edge by the ceaseless dash of the Atlantic. Last of all, apparently, went Anglesey, Wight, and the coastwise eyots, as well as the Bristol Channel. On the pro[75]tected eastern shore of Britain generally, the low slopes have survived well enough, and patches of shingle and sand, like the Dogger Bank, still mark the position of the higher sunken lands; but on the west and north the open Atlantic has eaten away all but the most sheltered plains, and cut its way at all exposed points into the heart of the hills, giving rise to the magnificent cliff scenery of Cornwall, Kerry, and the western Highlands. If you stand upon the shore of Coboe Bay in Guernsey, and look at low tide across the vast floor of jagged and water-fretted granite rocks which line its bottom, you will see with what force the waves have wormed their way over all the lowland; and they will only halt when they have planed down the whole of the island, as they have already planed down the lesser land which once stretched out to northward beyond the solitary pinnacles of the Casquets.
When all these changes had taken place, the stray members of the southern flora in Cornwall, Devon, Kerry, and Connemara would find themselves quite cut off from their fellows in the Mediterranean, the Pyrenees, and the Asturias. For the water has eaten away almost all the plain of the Bay of Biscay, save only a comparatively insignificant angle between the Loire, the mountains of Auvergne, and the roots of the[76] Pyrenees; and it has left the high and bleak granite moorland of Brittany jutting out alone into the western sea. But Brittany looks northward, and is open only to the chilliest winds; while its fair share of the Gulf Stream is diverted by currents due to the lay of the land in Cornwall. Moreover, the great bight of Biscay distracts and upsets the old run of the water, so that the whole shore of France from the Garonne northward is really colder and less equable in temperature than Cornwall and Kerry, or even than the average of our own western and southern coast. The Vendée is a chilly marshland; Bretagne Bretonannte is a high and wind-swept heath. On the other hand, our extreme south-western peninsulas and islands are bathed on every side by the warm water of the Gulf Stream, and so possess an unusually mild, damp, and equable climate. Every one has heard of the semi-tropical vegetation of palms and aloes which flourishes in the open air at Tresco Abbey in the Scilly Isles. Here, then, we have exactly the conditions under which the southern plants, though beaten back to the very base of the hills, might still manage to keep up a precarious existence in a few scattered and sheltered nooks. And that is exactly what they have done. Separated from all the rest of their kind, exposed to occasional hard winters or[77] heavy frosts, and slowly dying out under our very eyes, they have yet left here and there a few isolated descendants to tell the story of their origin and their failure. Curiously enough, these little lingering colonies of Mediterranean plants exist only on the southern and western slopes, among the cliffs and combes and bays which face and overlook the submerged lands whence their ancestors were driven by the advancing sea. So oddly do they confine themselves to the islands and the most insular peninsulas that their geographical distribution almost looks like a preconcerted arrangement.
Thus we may observe once more that one little islet of the Bristol Channel alone preserves the red pæony. Holyhead Island has half a dozen rare species. The Jersey centaury, Pelisser’s linaria, and several other southern flowers have died out everywhere save in the Channel Islands. Scilly shares with them in the sand bird’s foot. The Irish Arran and other Irish islands have many peculiar species; and a few southern types even reach Bute and the western Highlands; for, as every one knows, Rothesay has a climate almost as warm as Torquay. So, too, with the peninsulas. The Lizard, with the most equable temperature on the English coast, is a perfect mine of wealth to the botanist. It has three peculiar[78] southern clovers, and lots of other rarities. Penzance, at the very horn of Cornwall, has five or six specialities. The position of Kerry gives it a climate like that of Finisterre, with the appropriate flora. Wild madder belongs only to a few headlands of Pembrokeshire, the Damnonian peninsula, and the south-west of Ireland. Torquay, on the promontory of Hope’s Nose, shares a southern buplever with the Channel Islands. Babbicombe has a species almost to itself. Corfe Castle, in the so-called Isle of Purbeck in Dorset, divides a Spanish heather with Cornwall and the West of Ireland. One kind of rest-harrow, after getting up from the Pyrenees as far as the Channel Islands, then positively takes a second spring to the Mull of Galloway. As to the number of Mediterranean plants which are found in Britain only in Devon and Cornwall, or in Kerry and Connemara, or in both, I spare you the recital of them. Even the more inland and moorland types, which each survive on one high common alone, answer to the same law; for they occur on the warmest moors, in the neighbourhood of the sunniest south-western slopes. Thus the Cheddar pink grows in a single basking hollow heated by radiation from two great walls of limestone rock upon the western flanks of Mendip; the purple lobelia loiters on a bright[79] upland near the warm valley of the Devonshire Axe; the white sedum struggles on upon the edge of Malvern; and my hairy wood-spurge here battles hard for life on Claverton Down, close to the steaming basin of the old Roman Thermæ at Bath.
And so I end where I began. My sermon has led me far afield; but, like a good preacher, I have come back to my text. I have only touched lightly upon the simplest and least technical proofs; but when the whole evidence is put together—as I do not pretend to put it together off-hand, sitting here cross-legged on the edge of Wansdyke—there can be very little reasonable doubt that this is something like the way in which the hairy wood-spurge first found its way to the Prior Park Lane. So I have gathered my little morsel tenderly and carefully, not injuring the little plant more than I can help by my clumsiness; and I hope all future botanisers will do the same, in order to aid in preserving and handing down to after ages this interesting fragment of old English history, kept green and vital for us all in the tiny blossom of a wayside weed.

Fig. 17.—The Wild Strawberry.
Side by side in our English hedgerows in early springtime there grow two sister plants, almost exactly alike in foliage, flower, and all other points except the fruit, but differing widely from one another in that solitary, and to us essential, particular. One of these plants is the wild strawberry, the other is the little three leaved, white potentilla. It is not often that a parent species and its more developed offspring survive together in the same district, but this is almost certainly the case with these two small English wayside flowers. Indeed, the similarity between them is so close that even the most unobservant passers-by have been greatly struck with it; and the common native English name of the white potentilla—‘barren strawberry’—bears witness to the striking character of the family likeness. Perhaps one ought rather to go a step further, and to say that, while the most unobservant have perceived the relationship, only the more observant have ever discovered the distinctness[81] of the two plants. Nothing is more ordinary than to hear casual townsfolk exclaim that though there were lots of strawberry blossoms a little while ago in such-and-such a spot, there are no ripe strawberries to be seen now that the time has come for picking the fruit. In such cases, careful examination will generally show that the spot is really covered by white potentilla plants, whose little starry flowers were easily mistaken by the world at large for true straw[82]berry blossom. Though there are some marked distinctive features even in the flower, to which I shall presently recur, it is in the fruit alone that the two plants really differ sufficiently to attract the attention of an unbotanical eye. But here the difference is one which touches humanity on a very keen point indeed, for the strawberry blossom sets at last into a sweet and pulpy berry, while the potentilla blossom sets only into a small head of dry and unpalatable nutlets. How the edible fruit has developed from the inedible seeds is the question which I propose briefly to investigate in the present paper.
To get properly at the ancestry of the strawberry, we ought first to begin with the potentillas at large, for a most important part of our evidence consists in the fact that the white potentilla varies from the central type of its race in nearly all the same particulars as the strawberry plant. In other words, we have to show that the ancestors of the strawberry had already acquired most of their existing peculiarities while they were still white potentillas, and that they have only then varied so far as to have added to that white potentilla type the one extra peculiarity of a red and juicy berry. Our systematic botanists, indeed, will tell us that while the one plant belongs to the genus Potentilla, the other plant belongs to the[83] totally distinct genus Fragaria; and they imply, therefore, that the differences between the real strawberry and the barren strawberry are far greater than the differences between the barren strawberry and the other potentillas. I hope in the sequel to show, however, that it would be far easier to develop a strawberry out of a white potentilla than to develop a white potentilla itself out of any one among its yellow allies; and therefore that the systematic classification is a faulty one, and the popular classification a correct stroke of half-unconscious scientific intuition.
The potentillas are a group of very lowly and primitive roses, the earliest and simplest surviving members of the great and world-wide rose family. Our common English cinquefoil may be accepted as a good typical instance of the whole group. Cinquefoil is a pretty tufted creeping plant, whose small golden flowers, like yellow roses in miniature, star the waste grass-plots by the sides of lanes and highways everywhere in Britain during the summer and autumn months. Its leaves, as the very name denotes, consist of five separate spreading leaflets, all springing from a common point, and radiating round it as a centre like the fingers of a hand. The flowers, as usual in most very simple and primitive plants are bright golden yellow, and they closely resemble the[84] equally early blossoms of the buttercup, which similarly form the starting point of another great and varied family. Originally, there is good reason for believing, all flowers were of this same bright golden yellow hue; and those of them that have since progressed to other colours, under stress of special insect selection, have passed through regular gradations of white, pink, red, crimson, purple, and finally blue. Some flowers still remain at the ancestral yellow stage; others have got on as far as white or pink; yet others have attained the stage of crimson or purple; and a very few, the most advanced of all, have even reached the culminating glory of deep blue.
We have several other yellow potentillas in England besides the cinquefoil, and some of these have varied a good deal in foliage or other points from the central form. Nearest of all to it stands the small tormentil, so frequent upon heaths or other moors and uplands; for the main distinction between them lies in the fact that the cinquefoil has usually five large petals, while the tormentil has usually only four. This difference, however, is by no means always constant, for on the one hand it is easy to find stray flowers of cinquefoil with only four petals, while on the other hand the first flower on each stalk of tor[85]mentil has only five. There is an intermediate form, too, which exactly splits the difference between the two plants in every respect; and one can hardly doubt that tormentil is in reality only a very slightly altered form of cinquefoil, grown woodier and more dwarfish from its peculiar upland situation, and with one of its petals suppressed through gradual failure of constitutional vigour. The frequency with which the first flower on each stem recurs to the original five-petalled form, while the material to spare remains abundant, is very significant: the later flowers, as the material for their formation runs short, have generally to be content with only four petals each.
More divergent types of potentilla than these are the forms which have their leaves (to use the technical term) pinnately, not digitately, divided—that is to say, with the separate leaflets arranged along two sides of a central leaf-stalk instead of radiating from a common point; and though the white potentilla and the strawberry belong rather to the latter or digitate division, I shall yet enter briefly into the nature of the pinnate section, for the sake of the light which it throws by analogy upon the evolution of our own proper subject. Commonest among the potentillas of this divergent group in northern Europe is the trailing silver-weed or goose-weed of our English roadsides, a pretty, long[86]-leaved plant, with a silvery underside, and bright golden flowers springing from rooted joints on its creeping runners. A rarer plant is the shrubby potentilla, which grows in bushy or stony places, especially on mountain sides, and has accommodated itself to its special situation by acquiring a stout woody stem. This species also has a yellow flower. But there are two other pinnate-leaved English potentillas whose blossoms have long since changed colour under the selective influence of their insect fertilisers. One of these is the marshy comarum, a perennial which grows in peaty or boggy places, and has assumed a dingy purplish-yellow hue, to suit the eyes of marshland insects. It is very noticeable that waterside flies do not seem to care for yellow, and most waterside flowers are therefore pinkish, purplish, or white. Thus the water-crowfoot and the mud-haunting ivy-leaved crowfoot have become white, while all our other native buttercups remain yellow. In the group of bennets or Geums, closely allied to the potentillas, we find a still closer analogy, for the roadside herb-bennet or common avens is yellow like cinquefoil, but the marshy water-avens has exactly the same dusky purplish-yellow tint as the marshy comarum. The other pinnate English potentilla, found wild with us only among the clefts of the Breiddin[87] Hills in Montgomeryshire, is a mountain species with handsome and conspicuous white blossoms; and this also is in striking analogy with similar facts elsewhere, for mountain species usually rise higher than their neighbours in the scale of colour, owing to the keen competition between the flowers for the visits of those rare fertilisers, the butterflies, which sail further up mountain heights than the bees and other meadow honeysuckers. For example, some Alpine buttercups are snowy-white, while most of their lowland congeners are simply yellow.
With the side light thus cast upon our subject by the analogy of the pinnate potentillas, let us hark back to the digitate cinquefoil once more, and ask by what steps some such early ancestral form gave origin to the common predecessor of the true strawberry and its barren sister. The cinquefoil, we saw, had five leaflets to each leaf, but the strawberry and the white potentilla have three only. This is one of the marked points wherein these two plants differ from the other potentillas, and agree with one another. But though the trefoil leaf is a matter of some importance, as indicating community of origin, it is not difficult to understand how it has been developed from the primitive cinquefoil. The exact number of leaflets in a leaf is always rather variable, depending partly on the[88] mode of growth of the plant, and partly on the amount of available material. Thus, in the allied tormentil the lower leaves have five leaflets, but the upper ones have usually three only. In the spring potentilla, a rare English species, the lower leaves have seven or five, and the upper ones five or three. Again, where a species creeps along the ground, it is apt to have long pinnate leaves with many leaflets, as happens, for example, with silver-weed and many similar plants. But where the leaves grow habitually among tall grass or choking wayside weeds, the number of leaflets is very apt to be reduced to three, as happens, for example, with clover and lotus among the pea-flower tribe, and with wood-sorrel among the geranium tribe, many of whose allies have long pinnate leaves with numerous leaflets. Now, the strawberry and the barren strawberry differ conspicuously in habitat from the other potentillas in the fact that they grow mainly among grass, on banks, or in hedgerow thickets. Hence it suits them best to raise their trefoil leaves on tall stalks above the neighbouring herbage, and thus to get at the light and air which they require for their proper growth. Natural selection has easily brought about this result, because in such situations those potentillas which raised their leaves highest would best survive, while those which[89] trailed or crept closely along the ground would soon be starved out for want of carbonic acid (the raw material of growth) by their surrounding competitors.

| Fig. 18. Flower of Wild Strawberry. | Fig. 19. Flower of White Potentilla. |
In another direction the ancestors of the strawberry and of the barren strawberry also diverged from their cinquefoil predecessors, and that was in the peculiar colour of their flowers. For some reason rather difficult to decide, the petals have changed from yellow to white. Difficult to decide, I say, because we do not exactly know what are the insects which the strawberries set themselves out especially to please or what is the peculiar nature of their specific taste. But, as a rule, this change from yellow to white petals is an ordinary concomitant of higher development, and it probably accompanies some change in the insects to which fertilisation is generally due. Our own native species have got no further in the upward course of development than white; but two[90] allied East Indian forms with digitate leaves, cultivated in our flower-gardens, the Nepaul potentilla and the purple potentilla, have risen as far in the scale of coloration as crimson and deep red.

| Fig. 20. Fruit of Wild Strawberry. | Fig. 21. Fruit of White Potentilla. |
One may sum up the common points of the strawberry and the barren strawberry somewhat as follows: Both have tall leaves of three leaflets, raised on an elevated leaf-stalk, whereas most of their other congeners have many leaflets. Both have white flowers, whereas most of their other congeners have them yellow. Both have short tufted stems; in both the leaves are clothed with silky down; and in both the leaflets are regularly toothed at the edge in the self-same manner. On the other hand, they differ from one another almost exclusively in the matter of their fruit. Now, as we shall proceed to see, it is comparatively easy to produce the change whereby a dry fruit becomes a succulent one, and it is also compara[91]tively easy to produce any one single change unaccompanied by others; but it is comparatively difficult to produce the whole set of changes whereby the two strawberries differ alike from all their congeners. So, if we are going to make a new genus, Fragaria, with a Latin name at all, we ought to make it include both the true strawberry and the barren strawberry, while we relegate to the genus Potentilla all the other less closely related kinds. But perhaps we shall do better if we lump them all together in a single genus, considering that, after all, the barren strawberry does not differ more from the remainder of the potentillas than many of these differ from one another among themselves.
And now, how did the edible strawberry get developed from its barren ally? Well, if we take the fruit of any potentilla, we shall find that it consists of several small, dry, one-seeded nuts, so tiny that they look themselves like seeds, crowded on a thick receptacle or flower stalk, without any signs of redness or succulence. In some potentillas, however, as the fruit ripens, this receptacle becomes a little spongy, something like the hull of a raspberry, only without its pulpy character. It is a common tendency of fruits to develop such pulpiness, and sometimes they do so quite suddenly by apparently spontaneous variation, as when[92] an almond tree has been known to produce peach-like fruits. But no fruit will permanently acquire such a succulent character unless it derives some benefit by doing so: the change, once set up, will only be perpetuated by natural selection if it proves of advantage to the plants which happen to display it. Has it done so in the case of the strawberry?
A strawberry, as we all know, consists of a swollen red receptacle or end of the flower-stalk, dotted over with little seed-like nuts, which answer to the tiny dry fruits of the potentilla. Suppose any ancestral potentilla ever to have shown any marked tendency towards fleshiness in the berry, what would happen? It would probably be eaten by small hedgerow birds, who would swallow and digest the pulp, but would not digest the seed-like nuts embedded in its midst. Hence the nuts would get carried about from place to place and dropped by the birds in hedgerows or woods, under circumstances admirably adapted for their proper germination. Supposing this to happen often, the juiciest berries would get most frequently eaten, and so would produce hearty young plants oftener than those among their neighbours which simply trusted to dropping off casually among the herbage. Again, the birds like sweetness as well as pulpiness, and those berries which grew most full of sugary[93] juices would be most likely to attract their attention. Once more, the brightest-coloured fruits would be most easily seen among the tall foliage of the hedgerows, and so those berries which showed any tendency towards redness of flesh would be sure to gain a point in attractiveness over their greener rivals. Thus at last the strawberry has grown into the fruit that we know so well, by constant unconscious selection of the little hedgerow birds, exerted at once in favour of the pulpiest, the sweetest, and the ruddiest berries.
It is noticeable, too, that in all these particulars what happens to the strawberry happens also in a hundred other independent cases. Wherever animals are to be enticed by plants, sugar is sure to be developed to entice them. It is so developed in the honey of flowers, in the extra-floral nectaries used for attracting ants, and in the sweet secretion by which the pitcher plants allure flies into their murderous vessels. So, too, bright colour is commonly employed to advertise the sweet material, as in the petals of flowers, the skin of fruits, and the pink or purple patches on the lips of the pitcher plants. On the other hand, the particular way in which these allurements are displayed by the strawberry is very different from that adopted by almost all other fruits. In the closely allied raspberry, the pulpiness and[94] colour are produced in the outer coat of the little nutlets themselves, and the receptacle assumes the form of the hull, which we pull out of the fruit and throw away. In the plum, there is only one such berry, inclosing a single seed. But in the strawberry, the separate fruits remain always hard and dry, and it is only the receptacle which holds them that swells out into the bright-coloured and juicy edible portion.
It very seldom happens, however, that a plant which has diverged from another in one point remains constant in all other points. In the strawberry this is almost the case, for it hardly differs at all in any particular, save its fruit, from its ancestor, the white potentilla; and that is good evidence, it seems to me, that the two plants cannot very long have separated from one another. Yet even here there are a few inconspicuous lateral differences. Most notable of these are the variations in the flower. Though to a casual observer the two blossoms look almost identical, and the plants can only readily be identified when in fruit, a botanical eye has never any difficulty in distinguishing the one from the other. The petals of the barren strawberry are usually short and narrow, the flowers scarcely open into more than a cup shape, and there is a good deal of yellowish or reddish colour about the receptacle and the base of the[95] stamens. In the true wild strawberry, on the other hand, the petals are usually larger, rounder, and purer white, the flowers open into a wide saucer shape, and there is no yellow or red in the centre of the blossom. Perhaps one may best account for these changes by supposing that the true strawberry has still further progressed in insect fertilisation. This would sufficiently explain the purer white of the petals and the loss of such relics of the primitive yellow hue as still remained in the barren strawberry. But it is also probable, I think, that the barren strawberries represent the remnants of the old ancestral race which have not yet been lived down by the newer strawberry type, but which are gradually undergoing progressive degradation; hence their half-opened flowers—often self-fertilised—their smaller and degenerate petals, and their general unattractiveness of outward appearance. It is difficult to compare the blossom of a true wild strawberry with that of a barren strawberry without immediately catching the obvious suggestion that the one is going upward towards higher development and the other downward towards general degeneracy.
In some other respects the strawberry plant equally shows itself the nobler species of the two. Its leaves are usually larger and more erect, its stem taller and straighter, its root-stock less fluffy and[96] not so creeping. Moreover, if it really descended from the white potentilla, or from some closely allied common ancestor, it has certainly far outstripped its progenitor in the race for the possession of the world, for the white potentilla, or barren strawberry, is apparently a strictly European species, found from Sweden and Ireland to the Crimea and the Caucasus, but the true strawberry is a much more cosmopolitan plant, being found in almost all the temperate regions of the world, from Siberia and Scotland to Vancouver’s Land, and from the Arctic Regions to the Andes of Chili. This is quite what one would expect under the circumstances; for while the seed-like fruits of the white potentilla could only fall on the ground close to the mother plant, and so could disperse themselves very slowly over a single continent, the little nuts of the strawberry could be carried by birds from land to land, across the severing ocean or the intervening tropical region. Thus the old degenerate type is now apparently dying out in northern and western Europe; but the progressive and advancing strawberry is making its way steadily, like a colonising race, round the entire girdle of the two temperate regions.
The strawberries are, as yet, it would seem, a relatively new race, and so they have not, so far,[97] split up into any very marked or distinctive separate species. Still they have even now assumed several minor forms, worthy at least to be distinguished as nameable varieties. The most divergent of these, as might be expected, is the Chilian pine strawberry, for in the southern hemisphere the imported strawberry, carried over at first, no doubt, by some weather-driven bird, has found itself in the midst of a very different environment from that which surrounds it in the hedgerows and meadows of its European home: and to this environment it has had to adapt itself in several minor but obvious particulars. An almost equally distinct variety is the white Alpine strawberry, which has quite lost the native blushing ruddiness of the lowland fruit. Curiously different in another way is the hautboy, a taller plant, with fewer and larger blossoms and a richer flavour, chiefly distinguished by the separation of its sexes on distinct plants, for here the stamens grow on one vine, and the pistils, or embryo fruits, on another. In order to make the berries swell and ripen, it is necessary to plant both sorts together, and then the fertilising insects unconsciously carry the pollen from the staminate flowers to the sensitive surface of their pistillate neighbours, and so assist the efforts of the gardener in setting the fruit. In the great American market[98] gardens it is usual to plant one row of ‘barren’ flowers for every three rows of ‘fertile’ ones, leaving the insects to do the rest. At present none of these varieties can be said to have developed into what old-fashioned botanists used to call ‘a good species,’ for fertile cross-breeds can still be readily produced between them all by artificially fertilising the pistils of one sort with pollen taken on a camel’s-hair brush from the stamens of another. The possibility of fertile hybridisation in such a manner shows that the plants have not long diverged from the common central stock. But after they have long been exposed to varying circumstances and acted upon by natural selection, they will probably become so different from one another in a variety of small particulars that the hybrids will no longer prove fertile, owing to the want of sufficient similarity between the respective ancestral lines. Perfect fertility is only possible between individuals which still retain all the principal traits of a common ancestral form. Curiously enough, one existing variety, the Himalayan strawberry, has actually reverted to the primitive yellow flowers of its cinquefoil allies.
On the other hand, if the strawberries ever really live down the white potentillas, so that the latter race dies out altogether, then the distance between the[99] genus Fragaria and the genus Potentilla will be far greater than it is at the present day. We are lucky enough at this moment to be able to trace the close connection between one rather abnormal potentilla (the barren strawberry) and the true strawberry itself. But if the barren strawberry and the Himalayan kind were to disappear we should have no link between the yellow-flowered, five-leaved, dry-fruited cinquefoil and the white-flowered, three-leaved, succulent-fruited strawberry. In nature, as it now stands, the ‘missing link’ is fortunately not yet missing. Though still essentially a potentilla in all important points, it yet approaches so nearly to the true strawberry that only rather close observation enables us to perceive their differences in certain stages of their existence. What thus happens now with the strawberry has doubtless happened at one time or another with every new species of plant or animal; but the special interest of this case consists in the fact that here, in all probability, we still have the parent type living on in a degraded form side by side with its more advanced and developed descendant.
Fig. 22.—Goose-grass or Cleavers.
Sitting here on the gate that leads into the Fore Acre, I have just disentangled from my nether integuments a long trailing spray of clinging goose-grass, which has fastened itself to my legs by the innumerable little prickly hooks that line the angles of its four-cornered stem. It is well forward for the time of year, thanks to our wonderfully mild and genial winter; for it is already thickly covered with its tiny white star-shaped flowers, which have even set here and there into the final mature stage of small burr-like fruits. Goose-grass, or cleavers, as we ordinarily call it, is one of the very commonest among English weeds, and yet I dare say you never even heard its name till I told it to you just now; for it is an inconspicuous, petty sort of plant, which would never gain any attention at all if it were not for its rough clinging leaves, that catch one’s fingers slightly when drawn through them,[101] and often obtrude themselves casually upon one’s notice by looping themselves in graceful festoons about one’s person. Now I am glad to have got you button-holed here upon the gate, because I can tell you all about the goose-grass as we sit on the top bar without risk of interruption; and I dare say you will be quite surprised to learn that a very interesting and historical plant it is after all, in spite of its uninviting external aspect. You will find that its prickly leaves its square stem, its white flowers, and its odd little[102] fruit all tell us some curious incident in its past evolution, and are full of suggestiveness as to the general course of plant development. Here is our weed in abundance, growing all along the hedgerow by our side, and clambering for yards from its root over all the bushes and shrubs in the thicket. Pick a piece for yourself before I begin, and then you can follow my preaching at your leisure, with the text always open before you for reference and verification.
Fig 23.
Seedling of Cleavers.
Of course goose-grass had not always all its present marked peculiarities. Like every other living thing, it has acquired its existing shape by slow modification from a thousand widely different ancestral forms. One of the best ways to discover certain lost links in the pedigree of plants or animals is to watch the development of an individual specimen from the seed or the egg; for the individual, we have all often been told, to some extent recapitulates in itself the whole past history of its race. Thus the caterpillar shows us an early ancestral form of the butterfly, while it was still a wingless grub; and the tadpole shows us an early ancestral form of the frog, while it was still a limbless mud fish. So, too, the chick hatching within the shell goes through stages analogous to those of the fish, the amphibian, the reptile, and the bird successively. In just the same way young plants[103] pass through a first simple shape which helps us to picture to ourselves what they once were—what, for example, the ancestors of the goose-grass looked like, long before they were goose-grasses at all. Now here in my hand I have got a young specimen in its very earliest stage, which closely reproduces the primitive type of its first progenitors, a million ages since. Goose-grass is an annual weed: it dies down utterly every autumn, and only reproduces itself by seed in the succeeding spring; but this year the weather has been so exceptionally warm and summer-like that thousands of young plants have sprouted from the seed ever since Christmas; and among them is the specimen which I have just picked, and which you may have for examination if you will take the trouble. Look into it, and you will see that its two first leaves are quite unlike the upper ones—a phenomenon which frequently occurs in seedling plants, and with which you are probably familiar in the case of the pea and of the garden bean. But this difference is always a difference in one direction only; the first leaves which come out of the seed are invariably simpler in shape[104] and type than all the other leaves which come after them. In the language of science they are less specialised; they represent an earlier and undeveloped form of leaf—nature’s rough sketch, so to speak, while the later foliage represents the final improvements introduced with time, and perfected by the action of natural selection.
These large oval leaves which you see in the seedling are mere general models or central ideas of what a leaf should be; they are quite unadapted to any one special or definite situation. They are not divided into many little separate leaflets, or prolonged into points and angles, or gracefully vandyked round the edges, or beautifully cut out into lace-like patterns, or armed at every rib with stout defensive prickles, like many other leaves that you know familiarly. Their outline is quite simple and unbroken; they preserve for us still the extremely plain ancestral form from which such different leaves as those of the horse-chestnut, the oak, the clover, the milfoil, the parsley, and the holly are ultimately derived. An expanded oval, something like this, is the prime original, the central point from which every variety of foliage first set out, and from which they have all diverged in various directions, according as different circumstances favoured or checked their development[105] in this, that, or the other particular. Just as a single little cartilaginous mud-haunter—a blind and skulking small creature, something like a lancelet, something like a tadpole, and something like the famous ascidian larva—has gradually evolved, through diverse lines, all the existing races of beasts, birds, reptiles, and fishes, so too a single little primæval plant, something like these two lowest leaves of the goose-grass, has gradually evolved all the oaks and elms and ashes; all the roses, and geraniums, and carnations; all the cabbages, and melons, and apples, which we see in the world around us at the present day. And, again, just as the larval form of the ascidian and of the frog still preserves for us a general idea of that earliest ancestral vertebrate, so too these larval leaves of the goose-grass, if I may venture so to describe them, still preserve for us a general idea of that earliest dicotyledonous plant.
Dicotyledonous is a very ugly word, and I shall not stop now to explain it from the top of a five-barred gate. It must suffice if I tell you confidentially that the little plant we have ideally reconstructed was the first ancestor of almost all the forest trees, and of all the best known English herbs and flowers; but not of the lilies, the grasses, and the cereal kinds, which belong to the opposite or monocotyle[106]donous division of flowering plants. When this sprig of goose-grass first appeared above the ground, it probably represented that typical ancestor almost to the life; for it had then only the two rounded leaves you see at its base, and none of these six-rowed upper whorls, which are so strikingly different from them. Now, how did the upper whorls get there? Why, of course they grew, you say. Yes, no doubt, but what made them grow? Well, the first pair of leaves grew out of the seed, where the mother plant had laid by a little store of albumen on purpose to feed them, exactly as a reserve of food materials is laid by in the egg of a hen to feed the growing chick. Under the influence of heat and moisture the seed began to germinate, as we call it—that is to say, oxygen began to combine with its food stuffs, and motion or sprouting was the natural result. This motion takes in each plant a determinate course, dependent upon the intimate molecular structure of the seed itself; and so each seed reproduces a plant exactly like the parent, bar those small individual variations which are the ultimate basis of new species—the groundwork upon which natural selection incessantly works. In the case of this goose-grass seed the first thing to appear was the pair of little oval leaves; and, as the small store of albumen laid by in the seed was all used up in pro[107]ducing them, they had to set to work at once manufacturing new organic material for the further development of the plant. Luckily they happened to grow in a position where the sunlight could fall upon them—a good many seedlings are more unfortunate, and so starve to death at the very outset of their careers—and by the aid of the light they immediately began decomposing the carbonic acid of the air and laying by starch for the use of the younger generation of leaves. At the same time the vigorous young sap carried these fresh materials of growth into the tiny sprouting bud which lay between them, and rapidly unfolded it into such a shoot as you see now before you, with level whorls of quite differently shaped and highly developed leaves, disposed in rows of six or eight around the stem.
Observe that the adult type of leaf appears here suddenly and as it were by a leap. If we could reconstruct the whole past history of the goose-grass, we should doubtless find that each change in its foliage took place very gradually, by a thousand minute intermediate stages. Indeed, many of these stages still survive for us among allied plants. But the impulsive goose-grass itself clears the whole distance between the primitive ancestor and its own advanced type at a single bound. The intermediate[108] stages are all suppressed. This is not always the case: there are many plants which begin with a simple type of leaf, and gradually progress to a complex one by many small steps; just as the tadpole grows slowly to be a frog by budding out first one pair of legs and then another, and next losing his tail and his gills, and finally emerging on dry land a full-fledged amphibian. The goose-grass, however, rather resembles the butterfly, which passes at once from the creeping caterpillar to the complete winged form, all the intermediate stages being compressed into the short chrysalis period; only our plant has not even a chrysalis shape to pass through. It is in reality a very advanced and specially developed type—the analogue, if not of man among the animals, at least of a highly respectable chimpanzee or intelligent gorilla—and so it has learnt at last to pass straight from its embryo state as a two-leaved plantlet to its typical adult form as a trailing, whorled, and prickly creeper.
And now let us next look at this adult form itself. Here I have cut a little bit of it for you with my penknife, and, if you like, I will lend you my pocket lens to magnify it slightly. The fragment I have cut for you consists of a single half-inch of the stem, with one whorl of six long pointed leaves. You will[109] observe, first, that the stem is quadrangular, not round; secondly, that the leaves are lance-shaped, not oval; and thirdly, that both stem and leaves are edged with little sharp curved prickles, pointing backward the opposite way to the general growth of the plant. Let us try to find out what is the origin and meaning of these three marked peculiarities.
Fig. 24.—Stem of Cleavers.
To do so rightly we must begin by considering the near relations of the goose-grass. In a systematic botanical classification our plant is ranked as one of the stellate tribe, a subdivision of the great family of the Rubiaceæ, or madder kind. Now, the stellates are so called because of their little star-shaped flowers, and they are all characterised by two of these goose-grass peculiarities—namely, the square stems and the whorled leaves—while the third point, the possession of recurved prickles on the angles of the stalk and the edges of the leaves, is a special personal habit of the goose-grass species itself, with one or two more of its near relations. It will be best for us, therefore, to ask first what is the origin and meaning of the charac[110]teristics which our plant shares with all its tribe, and afterwards to pass on to those which are quite confined to its own little minor group of highly evolved species.
What, then, is the use to the goose-grass of these small, narrow, thickly whorled leaves? Why are they not all and always large, flat, and oval, like the two seed leaves? The answer must be sought in the common habits of all the stellate tribe. They are without exception small, creeping, weedy plants, which grow among the dense and matted vegetation of hedgerows, banks, heaths, thickets, and other very tangled places. Now, plants which live in such situations must necessarily have small or minutely subdivided leaves, like those of wild chervil, fool’s parsley, herb-Robert, and fumitory. The reason for this is clear enough. Leaves depend for their growth upon air and sunlight: they must be supplied with carbonic acid to assimilate, and solar rays to turn off the oxygen and build up the carbon into their system. In open fields or bare spaces, big leaves like burdock, or rhubarb, or coltsfoot can find food and space; but where carbonic acid is scarce, and light is intercepted by neighbouring plants, all the leaves must needs be fine and divided into almost thread-like segments. The competition for the carbon under such circum[111]stances is exceedingly fierce. For example, in water only very small quantities of gas are dissolved, so that all submerged water-plants have extremely thin waving filaments instead of flat blades; and one such plant, the water-crowfoot, has even two types of foliage on the same stem—submerged leaves of this lace-like character, together with large, expanded, floating leaves which loll upon the surface something like those of the water-lily. In the same way hedgerow weeds, which jostle thickly against one another, have a constant hard struggle for the carbon and the sunshine, and grow out accordingly into numerous small subdivided leaflets, often split up time after time into segments and sub-segments of the most intricate sort. I do not mean, of course, that each individual leaf has its shape wholly determined for it by the amount of sun and air which it in particular happens to obtain, but that each species has slowly acquired by natural selection the kind of leaf which best fitted its peculiar habitat. Those plants survive whose foliage adapts them to live in the circumstances where it has pleased nature to place them, and those plants die out without descendants whose constitution fails in any respect to square with that inconvenient conglomeration of external facts that we call their environment.
Fig. 25. Interpetiolar Stipules.
That is why the goose-grass and the other stellate weeds have foliage of this minute character, instead of broad blades like the two seed leaves. But all plants of tangly growth do not attain their end in precisely the same manner. Sometimes one plan succeeds best and sometimes another. In most cases the originally round and simple leaf gets split up by gradual steps into several smaller leaflets. In the stellate tribe, however, the same object is provided for in a widely different fashion. Instead of the primitive leaf dividing into numerous leaflets, a number of organs which were not originally leaves grow into exact structural and functional resemblance to those which were. Strictly speaking, in this whorl of six little lance-shaped blades, precisely similar to one another, only two opposite ones are true leaves; the other four are in fact, to use a very technical term, interpetiolar stipules. A stipule, you know, of course, is a little fringe or tag which often appears at the point where the leaf stalk joins the stem, and its chief use seems to be to prevent ants and other[113] destructive insects from creeping up the petiole. But in all the stellate plants the two little stipules on each side of each leaf have grown gradually out into active green foliar organs, to supplement and assist the leaves, until at last they have become as long and broad as the original leaflets, and have formed with them a perfect whorl of six or eight precisely similar blades. How do we know that? you ask. In this simple way, my dear sir. The other Rubiaceæ—that is to say, the remainder of the great family to which the stellate tribe belongs—have no whorls, but only two opposite leaves; and we have many reasons for supposing that they represent the simpler and more primitive type, from which the stellate plants are specialised and highly developed descendants. But between the opposite leaves grow a pair of small stipules, occupying just the same place as the whorled leaflets in the goose-grass; and in some intermediate species these stipules have begun to grow out into expanded green blades, thus preserving for us an early stage on the road towards the development of the true stellates. Accordingly, we are justified in believing that in the whorls of goose-grass the same process has been carried a step further, till leaves and stipules have at last become almost indistinguishable.
There is, however, one way in which we can still[114] distinguish the original true leaves of each whorl from the leaf-like stipules. Only two leaves out of the six ever have buds or branches proceeding from their axils; and this last token infallibly marks out for us which are the real primitive opposite pair, and which the spurious imitation.
What may be the use of the square stem it would be more difficult to decide. Perhaps it may serve to protect the plant from being trodden down and broken; perhaps by its angularity and stringiness it may render it unpalatable to herbivorous animals. This much at least is certain, that very few cows or donkeys will eat goose-grass. There is another large family of plants—the dead-nettle tribe—all of which have also square stems; and they are similarly rejected as fodder by cattle. Indeed, the very fact that the stellate tribe have become thus quadrangular, while the other and earlier members of the madder kind, like coffee and gardenia, have round stems, in itself suggests the idea that there must be some sufficient reason for the change, or else it would never have taken place; but, as in many other cases, what that reason may be I really cannot with any confidence inform you from my simple professional chair on the gate here. If I were only at Kew Gardens, now—well, that might be a different matter.
And now let us come down to the individual[115] peculiarities of the goose-grass, and ask what is the use of the wee recurved prickles which you can see thickly scattered on the stalk and whorls by the aid of my pocket lens. You observe that they occur all along each angle of the stem, and around the edge and midribs of the leaflets as well. If you try to pull a bit of goose-grass out of the thicket entire, you will soon see the function they subserve. The plant, you notice, resists your effort at once; the little prickles catch securely on to the bushes and defeat all endeavours to tear it away. It is these prickles, indeed, which are the raison d’être of the goose-grass as a separate species: they mark it off at once from almost all the other members of the same group. There are many allied kinds of galium in England (for galium is the botanical name of the genus), with very similar leaves and flowers, but they all grow in shorter bunches and frequent less thickly populated situations. Goose-grass, however, has survived and become a distinct kind just in virtue of these very hooks. By their aid it is enabled to scramble for many feet over hedges and bushes, though it is but an annual plant; and it thus makes use of the firm stem of yonder hawthorn and this privet bush by our sides to raise its leaves into open sunny situations which it could never reach with its own slender stalk[116] alone. Such an obvious improvement gives it an undoubted advantage in the struggle for life, and so in its own special positions it has fairly beaten all the other galiums out of the field. One of its common English names—Robin Run-the-hedge—sufficiently expresses the exact place in nature which it has thus adapted itself to fill and to adorn.
But how did the goose-grass first develop these little prickles? That is the question. Granting that their possession would give it an extra chance in the struggle for existence, if once they were to occur, how are we to account for their first beginning? In this way, as it seems to me. Viewed structurally, the stout little hooks which arm the stem and leaves are only thickened hairs. Now hairs, or long pointed projections from the epidermis, constantly occur in almost all plants, and in this very family they are found on the edges of the leaflets and on the angles of the stem among several allied species. But such hairs may easily happen to grow a little thicker or harder, by mere individual or constitutional variation; and in a plant with habits like the goose-grass every increase in thickness and hardness would prove beneficial, by helping the festoons to creep over the bushes among which they live. Thus generation after generation those incipient goose-grasses which best[117] succeeded in climbing would set most seed and produce most young, while the less successful would languish in the shade and never become the proud ancestors of future plantlets. Even the less highly developed species, such as the wall galium and the swamp galium, have little asperities on the edge of the stem; but, as they need to climb far less than the hedgerow goose-grass, their roughnesses hardly deserve to be described as prickles. Our own special subject, on the other hand, being a confirmed creeper, finds the prickles of immense use to it, and so has developed them to a very marked extent. The corn galium, too, which clings to the growing haulms or stubble of wheat, has learnt to produce very similar stout hooks; while the wild madder, which I suspect is far more closely related to goose-grass than many other plants artificially placed in the same genus, has prickles of like character, but much larger, by whose aid it trails over bushes and hedges for immense distances.
After the leaves and stem we have to consider the nature of the flower. Look at one of the blossoms on the piece I gave you, and you will easily understand the main points of its structure. You notice that it consists of a single united corolla, having four lobes joined at the base instead of distinct and separate petals, while the centre of course is occupied[118] by the usual little yellow knobs representing the stamens and pistil. Each goose-grass plant produces many hundreds of such flowers, springing in small loose bunches from the axils of the leaves. What we have to consider now is the origin and meaning of the parts which make them up.
Fig. 26.—Single flower of Cleavers.
We have already seen in dealing with the daisy that the really important organs of the blossom are the little central yellow knobs, which do all the active work of fertilising the ovary and producing the seeds. The stamens, as we then observed, manufacture the pollen, and when the pistil is impregnated with a grain of this golden dust the fruit begins to swell and ripen. But the corolla or coloured frill around the central organs, which alone is what we call a flower in ordinary parlance, shows that the goose-grass is one of those plants which owe their fertilisation to the friendly aid of insects. Blossoms of this sort usually seek to attract the obsequious bee or the thirsty butterfly by a drop of honey in their nectaries, supplemented by the advertising allurements of a sweet perfume and a set of coloured petals. So much knowledge about the functions of flowers in general[119] we have already acquired; the question for our present consideration is this: What gives the goose-grass flower in particular its peculiar shape, colour, and arrangement?
First of all, you will notice that it has a united corolla—a single fringe of bloom instead of several distinct flower leaves. This marks its position as a very proud one in the floral hierarchy; for you will remember that only the most advanced blossoms have their originally separate petals welded into a solid continuous piece. Once upon a time, indeed, the early ancestors of our little creeper had five distinct petals, like those of a dog-rose or a buttercup; but that was many, many generations since. In time these petals began to coalesce slightly at the base, so as to form a short tube as in the primrose; and, since this arrangement made it easier for the insect to fertilise the flowers, because he was more certain to brush his head in hunting for honey against the pollen-bearing stamens and the sensitive summit of the pistil, all the flowers which exhibited such a tendency gained a decided advantage over their competitors, and lived and flourished accordingly, while their less fortunate compeers went to the wall. So in the course of ages such tubular flowers, like harebells and heaths, became very common, and to a great[120] extent usurped all the best and most profitable situations in nature. Among them were the immediate ancestors of the goose-grass, which had then regular long tubular blossoms, instead of having a mere flat, disk-shaped corolla like the one you see in the goose-grass before you. But, for a reason which I will presently tell you, in the goose-grass tribe itself the tube has gradually become shorter and shorter again, till at last there is nothing left of it at all, and the corolla consists simply of four spreading lobes slightly joined together by a little rim or margin at the base.
How do we know, you ask, that the goose-grass is descended from such ancestral flowers having a long hollow tube? Why may it not be an early form of tubular blossom, a plant which is just acquiring such a type of flower, rather than one which has once possessed it and afterwards lost it? Well, my dear sir, your objection is natural; but we know it for this reason. I told you some time since that the other great branch of the madder family, which had stipules instead of whorled leaves, was thereby shown to be a more primitive form of the common type than the stellate tribe, in which these stipules have developed into full-grown leaves. Now, all these tropical madder-like plants have large tubular blossoms, perfectly developed; so that we may reasonably infer[121] the ancestors of the goose-grass had the same sort of flowers when they were at the same or some analogous stage of development. Moreover, amongst the stellate plants themselves there are several which still retain the long tubes to the blossom; and these are rather the less developed than the more developed members of the little group. Such are the pretty blue field-madder, which has a funnel-shaped corolla, and the sweet woodruff, which has bell-shaped flowers. But the galiums, which are the most advanced (or degraded) species of all, have the tube very short or hardly perceptible, and the more so in proportion as they are most widely divergent from the primitive type.
Fig. 27.
Flower of Field-madder.
Why, however, should a flower which was once tubular have lost its tube? If it was an advantage to acquire such a long narrow throat, must it not also be an advantage always to retain it? That depends entirely upon the nature of the circumstances to which the plant must adapt itself. Now the fact is, the original madder group seems to have had large and showy flowers, which were fertilised by regular honey-sucking insects, such as bees and butterflies and humming-bird hawk-moths. These are tropical shrubs,[122] often of considerable size, and of very different habits from our little goose-grass. But in the temperate regions, since the earth has begun to cool into zones, some of these rubiaceous plants have found out that they could get along better by becoming little creeping weeds; and these are the stellates, including our present friend. Accordingly they have mostly given up the attempt to attract big honey-sucking insects whose long proboscis can probe the recesses of jasmine or woodbine, and have laid themselves out to please the small flies and miscellaneous little beetles, which serve almost equally well to carry their pollen from head to head. Now the flowers which specially cater for such minor insects are usually quite flat, so that every kind alike can get at the honey or the pollen; and that, I fancy, is why the goose-grass and so many of its allies have lost their tubes. They are, in fact, somewhat degenerate forms, descended from highly adapted tropical types, but now readjusted to a humbler though more successful grade of existence.
Closely connected with this question is the other and very interesting problem of their colour. Why is goose-grass white? For the very same reason—because it wishes to attract all sorts of little insects impartially. For this purpose white is the best colour. Almost all flowers which thus depend for[123] fertilisation upon many different species of winged visitors are white. And, indeed, the sort of colour in each kind of stellate flower (as in all others) depends largely upon the sort of insects it wishes to attract. Thus the little field-madder, which has a long tube and is fertilised by honeysuckers of a high type, is blue or pink, as all the family once was, no doubt, before it began to bid for more vulgar aid. Then the lesser woodruff, or squinancy-wort, whose tube is shorter, has white cups tinged with lilac. The goose-grass and most of its neighbours, whose flowers have undergone still greater degeneration, are simply white, because they wish to please all parties equally, and white is of course the most neutral colour they could possibly assume. Finally, the lady’s bedstraw, which has no tube, depends upon little colour-loving beetles for fertilisation, and, like many other beetle flowers, it is bright yellow.
This order of degradation exactly reverses the upward order of chromatic progress; for, as flowers advance in type, they pass from yellow, which is the lowest colour, through white, pink, red, and lilac, to purple and blue, which are the highest. And when through any special cause they begin to retrogress, they pass backward through the same stages in inverse order.
Again, you may have observed that I said just now the primitive ancestor of the goose-grass had five petals. But the present united corolla has only four lobes instead of five, and it is this arrangement, apparently, which has gained for the whole tribe the name of stellate. Now the tropical Rubiaceæ, which we saw reason to believe represent an earlier stage of development than the goose-grass group, have usually five lobes to the corolla; and in this respect they agree in the lump with the whole great class of dicotyledonous plants to which they belong. Therefore we may fairly conclude that to have four lobes instead of five is a mark of further specialisation in the stellates; in other words, it is they that have lost a lobe, not the other madder-worts that have added one. This, then, gives us a further test of relative development—or perhaps we ought rather to say of relative degeneration—among the stellate tribe. Wild madder, whose flowers are comparatively large, has usually five lobes. Yellow crosswort has most of its blossoms four-lobed, interspersed with a few five-lobed specimens. Goose-grass occasionally produces large five-lobed flowers, but has normally only four lobes. The still smaller skulking species have almost invariably four only. In fact, the suppression of one original petal seems to be due to the general dwarfing[125] of the flower in most of the stellate tribe. The corolla has got too small to find room for five lobes, so it cuts the number down to four instead. This is a common result of extreme dwarfing. For example, the tiny central florets of the daisy ought properly to be pinked out into five points, representing the five primitive petals, but they often have the number reduced to four. So, too, in the little moschatel, the outer flowers of each bunch have five lobes, but the central one, which is crowded around and closely jammed by the others, has regularly lost one in every case.
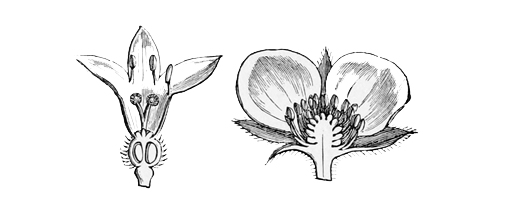
Fig. 28.—Strawberry and Asperula.
To show Inferior and Superior Ovaries.
There is just one more peculiarity of the goose-grass blossom which I must not wholly overlook. You see this rough little bulb or ball beneath the corolla, covered with incipient prickles? That is the part which will finally grow into the fruit, after some friendly insect has brought pollen on his legs from some neighbouring flower to impregnate the ovary of this. Now, what I want you to notice is the fact that the future fruit here lies below the corolla—below the flower, as most of us would say in ordinary language. But if you think of a strawberry, a raspberry, or a poppy, you will recollect that the part which is to become the fruit there grows above the corolla, and that the petals are inserted at its base. This last is[126] the original and normal position of the parts. How and why, then, has the ovary in the goose-grass kind managed to get below the petals? Well, the process has been something like this: When the flowers were tubular they were surrounded by a tubular calyx, and the ovary stood in the middle of both. But in the course of time, in order to increase the chances of successful fertilisation, the calyx tube, the corolla tube, and the ovary in the centre all coalesced into one solid piece—grew together, in fact, just as the five petals had already done. So now this little bulb really represents the calyx and ovary combined; while the corolla, only beginning to show at the top, where it expands into its four lobes, looks as if it started from the head of the fruit, whereas in reality it once started at the bottom, but has now so completely united with the calyx in its lower part as to[127] be quite indistinguishable. Thus the fruit is not in this plant a mere ripe form of the ovary, but is a compound organ consisting of the calyx outside, and the ovary inside, with the tube of the corolla quite crushed out of existence between them.

Fig. 29.
Fruit of Cleavers.
Last of all, let us look at the prickly fruit itself in its ripe condition. Some small fly has now fertilised the head with pollen from a brother blossom; the corolla and the stamens have fallen off; the embryo seeds within have begun to swell; the mother plant has stocked them with a little store of horny albumen to feed the tiny plantlets when they are first cast forth to shift for themselves in an unsympathetic world; and now the fruit here is almost ready to be detached from the stalk and borne to the spot where it must make its small experiment in getting on in life on its own account. Before I tell you how it manages to get itself transported free of cost to a suitable situation, I should like you to observe its shape and arrangement. It consists of two cells or carpels united in the middle, and each of these contains a single seed. Once upon a time there were several cells, as there still are in some of the tropical Rubiaceæ, and each cell contained several seeds, as is the case[128] with many of the southern species to the present day. But when the stellate tribe took to being small and weedy, they gave up their additional seeds and limited themselves to one only in each cell. This is another common result of the dwarfing process, and it is found again in all the daisy tribe and in the umbellates, such as fool’s parsley. To make up, however, for the loss in number of the seeds in each fruit, the number of fruits on each plant is still enormous. How many there are on a single weed of goose-grass I have never had the patience to count, but certainly not less than several hundred. You might find it a nice amusement for a statistical mind to fill up this lacuna in our botanical knowledge.
Most of the stellate plants have simple little fruits without any special means of dispersion, but in the goose-grass the same sort of prickles as those of the stem and leaves are further utilised for carrying the seed to its proper place. You know seeds have many devices for ensuring their dispersion to a distance from the mother plant. Some are surrounded by edible pulp, as in the case of the raspberry or the gooseberry; and these are swallowed by birds or animals, through whose body they pass undigested, and thus get deposited under circumstances peculiarly favourable to their germination and growth. Others[129] have little wings or filaments, as in the case of the dandelion or the valerian; and these get blown by the wind to their final resting-place. Yet others, again, are provided with hooks or prickles, like the burr and the hounds-tongue, by whose means they cling to the wool of sheep, the feathers and legs of birds, or the hair of animals, and thus get carried from hedge to hedge and rubbed off against the bushes, so as to fall on to the ground beneath. Now this last plan is especially well adapted for a plant like the goose-grass, which lives by straggling over low brambles and hawthorns, for it ensures the deposition of the seed in the exact place where the full-grown weed will find such support and friendly assistance as it peculiarly requires. Accordingly, we may be sure that if any half-developed goose-grass ever showed any tendency to prickliness on its fruit, it would gain a great advantage over its neighbours in the struggle for existence, and the tendency would soon harden under the influence of natural selection into a fixed habit of the species. Is there any way in which such a tendency could be set up?
Yes, easily enough, as it seems to me. You remember the outer coat of the fruit is really the calyx, and this calyx would be naturally more or less hairy, like the original leaves. We have only to sup[130]pose that the calyx hairs followed suit with the stem hairs, and began to develop into stiff prickles, in order to understand how the burr-like mechanism was first set up. Supposing it once begun, in ever so slight a degree, every little burr which succeeded in sticking to a sheep’s legs or a small bird’s breast would be pretty sure, sooner or later, of reaching a place where its seeds could live and thrive. It is from this habit of cleaving or sticking to one’s legs that the plant has obtained one of its English names—cleavers. Moreover, to make the development of the burr all the more comprehensible, many of the other galiums have rather rough or granulated fruits, while one kind—the wall galium—which in England has smooth or warty fruit, has its surface covered in southern Europe with stiff hairs or bristles. Another English galium besides goose-grass has hooked bristles on its fruit, though they are not so hard or adhesive as in our own proper subject. Thus the very steps in the evolution of the bristly fruit are clearly preserved for us to the present day in one or other of the allied species.
On the other hand, the very similar little corn galium, which has prickles on its stem and leaves to enable it to cling to the growing straw in the wheat-fields, has no hooks at all upon its fruit. Instead of a burr it produces only little rough-looking knobs or[131] capsules. At first sight, this difference between the plants is rather puzzling, but when we come to consider the peculiar habits of the corn galium we can see at once the reason for the change. Like most other cornfield weeds, it blossoms with the wheat, and its seed ripens with the mellowing of the shocks. Both are cut down together, and the seed of the galium is thrashed out at the same time as the grain. Thus it gets sown with the seed corn from year to year, and it would only lose by having a prickly fruit, which would get carried away to places less adapted for its special habits than the arable fields. It has accommodated itself to its own peculiar corner in nature, just as the goose-grass has accommodated itself to the hedgerows and thickets. So again, in the wild madder, the fruit, instead of becoming rough and clinging, has grown soft and pulpy, so as to form a small blackish berry, much appreciated by birds, who thus help unconsciously to disperse its seeds. Each plant simply goes in the way that circumstances lead it, and that is why we get such infinite variety of detail and special adaptation even within the narrow limits of a single small group.
And now I think you are tired both of your seat on the gate and of my long sermon. Yet the points to which I have called your attention are really only[132] a very few out of all the facts which go to make up the strange, eventful life-history of this little creeper. If you had but leisure and patience to hear me I might go on to point out many other curious details of organisation which help us to reconstruct the family pedigree of the goose-grass. There is not a single organ in the plant which does not imply whole volumes of unwritten ancestral annals; and to set them all forth in full would require not a single hour but a whole course of ten or twenty sermons. Still, I hope I have done enough to suggest to you the immense wealth of thought which the goose-grass is capable of calling up in the mind of the evolutionary botanist; and I trust, when you next get your clothes covered with those horrid little cleavers, you will be disposed to think more tenderly and respectfully than formerly of an ancient and highly developed English weed.
Wheat ranks by descent as a degenerate and degraded lily. Such in brief is the text which this paper sets out to prove, and which the whole course of evolutionary botany tends every day more and more fully to confirm. By thus from the very outset placing clearly before our eyes the goal of our argument, we shall be able the better to understand as we go whither each item of the cumulative evidence is really tending. We must endeavour to start with the simplest forms of the great group of plants to which the cereals and the other grasses belong, and we must try to see by what steps this primitive type gave birth, first to the brilliantly coloured lilies, next to the degraded rushes and sedges, and then to the still more degenerate grasses, from one or other of whose richer grains man has finally developed his wheat, his rice, his millet, and his barley. We shall thus trace throughout the whole pedigree of wheat from the time when[134] its ancestors first diverged from the common stock of the lilies and the water-plantains, to the time when savage man found it growing wild among the untilled plains of prehistoric Asia, and took it under his special protection in the little garden plots around his wattled hut, whence it has gradually altered under his constant selection into the golden grain that now covers half the lowland tilth of Europe and America. There is no page in botanical history more full of genuine romance than this; and there is no page in which the evidence is clearer or more convincing for those who will take the easy trouble to read it aright.
Fig. 30.
Head of Wheat.
Moreover, the case of wheat is a very interesting one, after the case of the daisy and of cleavers, because it exhibits a different order of evolution, that namely of continuous degradation. While the daisy has gone constantly up, and while the goose-grass has fallen but a little after a long course of upward development, the grasses generally have from the very first exhibited a constant and unbroken structural decline.
The fixed point from which we start on our[135] inquiry is the primitive and undifferentiated ancestral flowering plant. Into the previous history of the line from which the cereals are ultimately descended, I do not propose here to enter. It must suffice for our present purpose to say dogmatically that the flowering plants as a whole derive their origin from a still earlier flowerless stock, akin in many points to the ferns and the club-mosses, but differing from them in the relatively important part borne in its economy by the mechanism for cross-fertilisation. The earliest flowering plant of the great monocotyledonous division (the only one with which we shall here have anything to do) started apparently by possessing a very simple and inconspicuous blossom, with a central row of three ovaries, surrounded by two or more rows of three stamens each, without any coloured petals or other ornamental adjuncts of any sort. I need hardly here explain even to the unbotanical reader that the ovaries contain the embryo seeds, and that they only swell into fertile fruits after they have been duly impregnated by pollen from the stamens, preferably those of another plant, or at least of another blossom on the same stem. Seeds fertilised by pollen from their own flower, as Mr. Darwin has shown, produce relatively weak and sickly seedlings; seeds fertilised by pollen from a sister plant of the same species[136] produce relatively strong and hearty seedlings. The two cases are exactly analogous to the effects of breeding in and in or of an infusion of fresh blood among races of men and animals. Hence it naturally happens that those plants whose organisation in any way favours the ready transference of pollen from one flower to another gain an advantage in the struggle for existence, and so tend on the average to thrive and to survive; while those plants whose organisation renders such transference difficult or impossible stand at a constant disadvantage in the race for life, and are liable to fall behind in the contest, or at least to survive only in the most unfavourable and least occupied parts of the vegetal economy. Familiar as this principle has now become to all scientific biologists, it is yet so absolutely necessary for the comprehension of the present question, whose key-note it forms, that I shall make no apology for thus once more stating it at the outset as the general law which must guide us through all the intricacies of the development of wheat.
Our primitive ancestral lily, not yet a lily or anything else nameable in our existing terms, had thus to start with, one triple set of ovaries, and about three triple sets of pollen-bearing stamens; and to the very end this triple arrangement may be traced under more or less difficult disguises in every one of its numerous[137] modern descendants. It thus differed from the primitive ancestor of dicotyledonous flowers like the daisy and the goose-grass, which as we have seen had its parts arranged in whorls of five, not in whorls of three, like the ancestral lily. No single survivor, however, now represents for us this earliest ideal stage; we can only infer its existence from the diverse forms assumed by its various divergent modifications at the present day, all of which show many signs of being ultimately derived from some such primordial and simple ancestor. The first step in advance consisted in the acquisition of petals, which are now possessed in a more or less rudimentary shape by all the tribe of trinary flowers, or at least if quite absent are shown to have been once present by intermediate links or by abortive rudiments. There are even now flowers of this class which do not at present possess any observable petals at all; but these can be shown (as we shall see hereafter) not to be unaltered descendants of the prime type, but on the contrary to be very degraded and profoundly modified forms, derived from later petal-bearing ancestors, and still connected with their petal-bearing allies by all stages of intervening degeneracy. The original petalless lily has long since died out before the fierce competition of its own more advanced[138] descendants; and the existing petalless reeds or cuckoo-pints, as well as the apparently petalless wheats and grasses, are special adaptive forms of the newer petal-bearing rushes and lilies.
The origin of the coloured petals, we know, is almost certainly due to the selective action of primæval insects. The soft pollen, and perhaps too the slight natural exudations around the early flowers, afforded food to the ancestral creatures not then fully developed into anything that we could distinctively call a bee or a butterfly. But as the insects flew about from one head to another in search of such food, they carried small quantities of pollen with them from flower to flower. This pollen, brushed from their bodies on to the sensitive surface of the ovaries, fertilised the embryo seeds, and so gave the fortunate plants which happened to attract the insects all the benefits of a salutary cross. Accordingly, the more the flowers succeeded in attracting the eyes of their winged guests, the better were they likely to succeed in the struggle for existence. In some cases, the outer row of stamens appears to have become flattened and petal-like, as still often happens with plants in the rich soil of our gardens; and in these flatter stamens the oxidised juices assumed perhaps a livelier yellow than even the central stamens them[139]selves. If the flowers had fertilised their own ovaries this change would of course have proved disadvantageous, by depriving them entirely of the services of one row of stamens; for the new flattened and petal-like structures lost at once the habit of producing pollen. But their value as attractive organs for alluring the eyes of insects more than counterbalanced this slight apparent disadvantage; and the new petal-bearing blossoms soon outstripped and utterly lived down all their simpler petalless allies. By devoting one outer row of stamens to the function of alluring the fertilising flies, they have secured the great benefit of perpetual cross fertilisation, and so have got the better of all their less developed competitors. At the same time, the exudations at the base of the petals have assumed the definite form of sweet nectar or honey, a liquid which is mainly composed of sugar, that universal allurer of animal tastes. By this means the plants save their pollen from depredations, and at the same time offer the insects a more effectual because a more palatable sort of bribe.
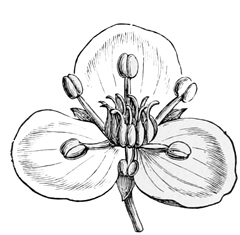
Fig. 31.—Single flower of Alisma plantago.
Passing rapidly over these already familiar initial stages, we may go on to those more special and distinctive facts which peculiarly concern the ancestry of the lilies and cereals. It is probable that the nearest modern analogue of the earliest petal-bearing trinary[140] flowers is to be found in the existing alisma tribe, including our own English arrowheads and flowering rushes. As a rule, indeed, it may be said that freshwater plants and animals tend to preserve for us very ancient types indeed; and all the alismas are marsh or pond flowers of an extremely simple character. They have usually three greenish sepals outside each blossom, inclosing one whorl of three white or pink petals, two or three whorls of three stamens each, and a number of separate ovaries, which are not united, as in the more developed true lilies, into a single capsule, but remain quite distinct, each with its own individual stigma or sensitive surface. Even within this relatively early and simple group, however, several gradations of development may yet be traced. I incline to believe that our English smaller alisma, a not uncommon plant in wet ditches and marshes throughout[141] the whole of southern Britain, represents the very earliest petal-bearing type in this line of development; indeed, save that its petals are now pinky-white, while those of the original ancestor were almost certainly yellow, we might almost say that the marsh-weed in question was really the earliest petal-bearing plant of which we are in search. It closely resembles in appearance, and in the arrangement of its parts, the buttercups, which are the earliest existing members of the other or quinary division of flowering plants; and in both we seem to get a survival of a still earlier common ancestor, only that in the one the parts are arranged in rows of three, while in the other they are arranged in rows of five; and concomitantly with this distinction go the two or three other distinctions which mark off the two main classes from one another—namely, that the one has leaves with parallel veins, only one seed-leaf to the embryo, and an endogenous stem, while the other has leaves with netted veins, two seed-leaves to the embryo, and an exogenous stem. Nevertheless, in spite of such fundamental differences, we may say that the alismas and the buttercups really stand very close to one another in the order of development. When the two main branches of flowering plants first diverged from one another, the earliest petal-bearing form they produced[142] on one divergent branch was the alisma, or something very like it; the earliest petal-bearing form they produced on the other divergent branch was the buttercup, or something very like it. Hence, whenever we have to deal with the pedigree of either great line, the fixed historical point from which we must needs set out must always be the typical alismas or the typical buttercups. The accompanying diagram will show at once the relation of parts in the simplest trinary flowers, and will serve for comparison at a later stage of our argument with the arrangement of their degraded descendants, the wheats and grasses.
a, ovaries; b, stamens, inner whorl; c, stamens, outer whorl; d, petals; e, calyx-pieces.
Fig. 32.—Diagram of primitive monocotyledonous flower.
Our own smaller alisma has a number of ovaries loosely scattered about in its centre, as in the butter[143]cups, with two rows of three stamens outside them, and then a single row of three petals, followed by the calyx or inclosing cup of three green pieces. Its close ally the water-plantain, however, shows signs of some advance towards the typical lily form in the arrangement of its ovaries in a single ring, often loosely divisible into three sets. And in the pretty pink flowering rush (not of course a rush at all in the scientific sense) the advance is still more marked in that the number of ovaries is reduced to six, that is to say, two whorls of three each, accompanied by nine stamens, similarly divisible into three rows. In all these very early forms (as in their analogues the buttercups) the main point to notice is this, that there is as yet no regular definiteness in the numerical relations of the parts. They tend to run, it is true, in rows of three; but often these rows are so numerous and so confused that nature loses count, so to speak, and it is only in their higher and more developed members that we begin to arrive at any distinct symmetry, such as that of the flowering rush. Even here, the symmetry is far from being so perfect as in the later lilies. There are, however, a few very special members of the alisma family in which the approach to the true lilies is even greater. These are well represented in England by our own common arrow[144]-grasses—inconspicuous little green flowers, with three calyx-pieces, three petals, six stamens, and either six or three ovaries. Here, too, the ovaries are at first united into a single pistil (as it is technically called), though they afterwards separate as they ripen into three or six distinct little capsules. One of our British kinds, the marsh arrow-grass, has almost reached the lily stage of development; for it has three calyx-pieces, three petals, six stamens, and three ovaries, exactly like the true lilies; but it falls short of their full type in the fact that its pistil divides when ripe into separate capsules, whereas the pistil of the lilies always remains united to the very end; and this minute difference suffices, in the eyes of systematic botanists, to make it an alisma rather than a lily. In reality, it ought to be regarded as a benevolent neutral—a surviving intermediate link between the two larger classes.
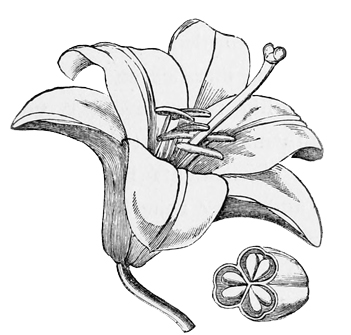
Fig. 33.—Flower of White Lily and section of ovary.
The specialisation which makes the true lilies thus depends upon two points. In the first place, all the parts are regularly symmetrical, except that there are two rows of stamens to each one of the other organs: the common formula being three calyx-pieces, three petals, six stamens, and three ovaries. In the second place, the three ovaries are completely combined together into a single three-celled pistil. The advan[145]tage which the lilies thus gain is obvious enough. Their bright petals, usually larger and more attractive than those of the alismas, allure a sufficient number of insects to enable them to dispense with the numerous stamens and ovaries of their primitive ancestors. Moreover, this diminution in number is accompanied by an increase in effectiveness and specialisation: for the lilies have only three sensitive surfaces to their pistil, combined on a single stalk: and the honey is usually so placed at its base that the insect cannot fail to brush off pollen at every visit against all three surfaces at once. Again, while the number of ovaries has been lessened, the number of seeds in each has been generally increased, which also[146] marks a step in advance, since it allows many seeds to be impregnated by a single act of pollination. The result of all these improvements, carried further by some lilies than by others, is that the family has absolutely outstripped all others of the trinary class in the race for the possession of the earth, and has now occupied all the most favourable positions in every part of the world. While the alismas and their allies have been so crowded out that they now linger only in a few ponds, marshes, and swamps, to which the more recent lily tribe have not yet had time fully to adapt themselves, the true lilies and their yet more advanced descendants have taken seizin of every climate and every zone upon our planet, and are to be found in every possible position, from the arborescent yuccas and huge agaves of the tropics to the wild hyacinths of our English woodlands and the graceful asphodels of the Mediterranean hill-sides.
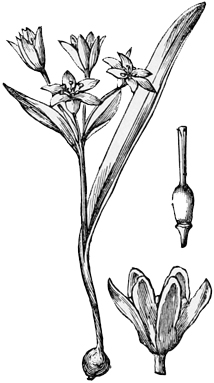
Fig. 34.—Gagea lutea.
The lilies themselves, again, do not all stand on one plane of homogeneous evolution. There are different grades of development still surviving among the class itself. The little yellow gagea (fig. 34) which grows sparingly in sandy English fields may be taken as a very fair representative of the simplest and earliest true lily type. It bears a small bunch of little golden[147] flowers, only to be distinguished from the higher alismas by their united ovaries: for though both calyx and petals are here brightly coloured, that is also the case in the flowering rushes, and in many others of the alisma group. On the other hand, though it may be said generally of the lilies that their calyx and petals are coloured alike—sometimes so much so as to be practically indistinguishable—yet there are many kinds which still retain the greenish calyx-pieces, and that even in the more developed genera. But most of the lilies are far handsomer than gagea and its allies: even in England itself we have such very conspicuous and attractive flowers as the purple fritillaries, which every Oxford man has gathered by handfuls in the spongy meadows about Iffley lock, with their dark spotted petals converging into a bell, and the nectaries at the base producing each a large drop of luscious honey. Some, like our wild hyacinths, have assumed a tubular shape under stress of insect selection, the[148] better to promote proper fertilisation; and at the same time have acquired a blue pigment, to allure the eyes of azure-loving bees. Others have become dappled with spots to act as honey-guides, or have produced brilliant variegated blossoms to attract the attention of great tropical insects. Our British lilies alone comprise such various examples as the lily-of-the-valley, a tubular white scented species, adapted for fertilisation by moths; the very similar Solomon’s seal; the butcher’s broom; the wild tulip; the star-of-Bethlehem; the various squills; the asparagus; the grape hyacinth; and the meadow saffron. Some of them (for example, asparagus and butcher’s broom) have also developed berries in place of dry capsules; and these berries, being eaten by birds which digest the pulp, but not the actual seeds, aid in the dispersion of the seedlings, and so enable the plant to reduce the total number of seeds to three only, or one in each ovary. Among familiar exotics of the same family may be mentioned the hyacinth, tuberose, tulip, asphodel, yucca, and most of the so-called lilies. In short, no tribe supplies us with a greater number of handsome garden flowers, for the most part highly adapted to a very advanced type of insect fertilisation.
Properly to understand the development of our[149] existing wheat from this brilliant and ornamental family, as well as to realise the true nature of its relation to allied orders, we must first glance briefly at the upward evolution of the other branches descended from the true lilies, and then recur to the downward evolution which finally resulted in the production of the degenerate grasses. In the main line of progressive development, the lilies gave origin to the amaryllids, familiarly represented in England by the snowdrops and daffodils, a family which is technically described as differing from the lilies in having an inferior instead of a superior ovary—that is to say, with the pistil apparently placed below instead of above the point where the petals and calyx-pieces are inserted. From the evolutionary point of view, however, this difference (as we saw in the case of the goose-grasses) merely amounts to saying that the amaryllids are tubular lilies, in which the tube has coalesced with the walls of the ovary, so that the petals seem to begin at its summit instead of at its base. The change gives still greater certainty of impregnation, and therefore benefits the race accordingly. At the same time, the amaryllids, being probably a much newer development than the true lilies, have not yet had leisure to gain quite so firm a footing in the world; though on the other hand many[150] of them are far more minutely adapted for special insect fertilisation than their earlier allies. They include the so-called Guernsey lilies of our gardens, as well as the huge American aloes which all visitors to the Riviera know so well on the dry hills around Nice and Cannes. The iris family are a similar but rather more advanced tribe, with only three stamens instead of six, their superior organisation allowing them readily to dispense with half their complement, and so to attain the perfect trinary symmetry of three sepals, three petals, three stamens, and three ovaries. Among them, the iris and the crocus are circular in shape, but some very advanced types, such as the gladiolus, have acquired a bilateral form, in correlation with special insect visits. From these, the step is not great to the orchids, undoubtedly the highest of all the trinary flowers, with the triple arrangement almost entirely obscured, and with the most extraordinary varieties of adaptation to fertilisation by bees or even by humming-birds in the most marvellous fashions. Alike by their inferior ovary, their bilateral shape, their single stamen, their remarkable forms, their brilliant colours, and their occasional mimicry of insect life, the orchids show themselves to be by far the highest of the trinary flowers, if not, indeed, of the entire vegetable world.
From this brief sketch of the main line of upward evolution from lilies to orchids, we must now return to the grand junction afforded us by the lilies themselves, and travel down the other line of degeneracy and degradation which leads us on to the grasses and the cereals, including at last our own familiar cultivated wheat. Any trinary flower with three calyx-pieces, three petals, six stamens, and a three-celled pistil not concealed within an inclosing tube, is said to be a lily, as long as it possesses brightly coloured and delicate petals. There are, however, a large number of somewhat specialised lilies with very small and inconspicuous petals, which have been artificially separated by botanists as the rush family, not because they were really different in any important point of structure from the acknowledged lilies, but merely because they had not got such brilliant and handsome blossoms. These despised and neglected plants, however, supply us with the first downward step on the path of degeneracy which leads at last to the grasses, and they may be considered as intermediate stages in the scale of degradation, fortunately preserved for us by exceptional circumstances to the present day. Even among the true lilies, there are some, like the garlic and onion tribe, which show considerable marks of degeneration, owing to some decline from the type[152] of insect fertilisation to the undesirable habit of fertilising themselves. Thus, while our common English rampsons or wild garlic has pretty and conspicuous white blossoms, some other members of the tribe, such as the crow allium, have very small greenish flowers, often reduced to mere shapeless bulbs. Among the true rushes, however, the course of development has been somewhat different. These water-weeds have acquired the habit of trusting for fertilisation to the wind, which carries the pollen of one blossom to the sensitive surface of another, perhaps at less trouble and expense to the parent plant than would be necessary for the allurement of bees or flies by all the bribes of brilliant petals and honeyed secretions. To effect this object, their stamens hang out pensile to the breeze, on long slender filaments, so lightly poised that the merest breath of air amply suffices to dislodge the pollen: while the sensitive surface of the ovaries is prolonged into a branched and feathery process, seen under the microscope to be studded with adhesive glandular knobs, which readily catch and retain every golden grain of the fertilising powder which may chance to be wafted toward them on the wings of the wind. Under such circumstances, the rush kind could only lose by possessing brightly coloured and attractive[153] petals, which would induce insects uselessly to plunder their precious stores: and so all those rushes which showed any tendency in that direction would soon be weeded out by natural selection; while those which produced only dry and inconspicuous petals would become the parents of future generations, and would hand on their own peculiarities to their descendants after them. Thus the existing rushes are all plain little lilies with dry brownish flowers, specially adapted to wind-fertilisation alone.
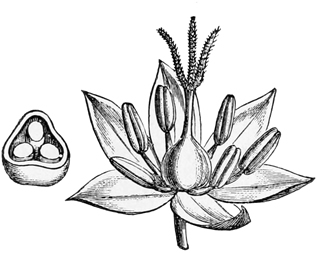
Fig. 35.—Single flower of Woodrush.
Among the rushes themselves, again, there are various levels of retrogressive development—retrogressive, that is to say, if we regard the lily family as an absolute standard; for the various alterations undergone by the different flowers are themselves adaptive to their new condition, though that condition is itself decidedly lower than the one from which they started. The common rush and its immediate congeners resemble the lilies from which they spring in having several seeds in each of the three cells which compose their pistil. But there is an interesting group of small grass-like plants, known as wood-rushes, which combine all the technical characteristics of the true rushes with a general character extremely like that of the grasses. They have long, thin, grass-like blades in the place of leaves; and what is still[154] more important, as indicating an approach to the essentially one-seeded grass tribe, they have only three seeds in the flower, one to each cell of the capsule. These seeds are comparatively large, and are richly stored with food-stuffs for the supply of the young plantlet. One such richly supplied embryo is worth many little unsupported grains, since it stands a much better chance than they do of surviving in the struggle for existence. The wood-rushes may thus be regarded as some of the earliest plants among the great trinary class to adopt those tactics of storing gluten, starch, and other food-stuffs along with the embryo, which have given the cereals their acknowledged superiority as producers of human food. They are closely connected with the rushes, on the one hand, by sundry intermediate species which possess thin leaves instead of cylindrical pithy blades;[155] and they lead on to the grasses, on the other, by reason of their very grass-like foliage, and their reduced number of large, well-furnished, starchy seeds.
In another particular, the rush family supplies us with a useful hint in tracing out the pedigree of the grasses and cereals. Their flowers are for the most part crowded together in large tufts or heads, each containing a considerable number of minute separate blossoms. Even among the true lilies we find some cases of such crowding in the hyacinths and the squills, or still better in the onion and garlic tribe. But with the wind-fertilised rushes, the grouping together of the flowers has important advantages, because it enables the pollen more easily to fix upon one or other of the sensitive surfaces, as the stalks sway backward and forward before a gentle breeze. Among yet more developed or degraded wind-fertilised plants, this crowding of the blossoms becomes even more conspicuous. A common American rush-like water-plant, known as eriocaulon, helps us to bridge over the gap between the rushes and such compound flowers as the sedges and grasses. Eriocaulon and its allies have always one seed only in each cell of the pistil: and they have also generally a very delicate corolla and calyx, of from four to six pieces, representing the original three sepals and three petals[156] of the lilies and rushes. But their minute blossoms are closely crowded together in globular heads, the stamens and pistils being here divided in separate flowers, though both kinds of flowers are combined in each head. From an ancestral form not unlike this, but still more like the wood rushes, we must get both our sedges and our grasses. And though the sedges themselves do not stand in the direct line of descent to wheat and the other cereals, they are yet so valuable as an illustration from their points of analogy and of difference that we must turn aside for a moment to examine the gradual course of their evolution.
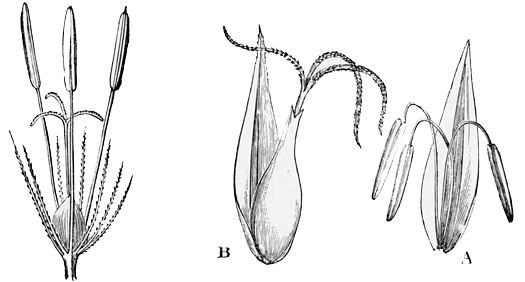
| Fig. 36. | Fig. 37. |
| Single flower of Scirpus. | Male and Female flower of Carex. |
The simplest and most primitive sedges now surviving, though very degenerate in type, yet retain some distinct traces of their derivation from earlier rush-like and lily-like ancestors. In the earliest existing type, known as scirpus, the calyx and petals which were brightly coloured in the lilies, and which were reduced to six brown scales in the rushes, have undergone a further degradation to the form of six small dry bristles, which now merely remain as rudimentary relics of a once useful and beautiful structure. In some species of scirpus, too, the number of these bristles is reduced from six to four or three. There is still one whorl of three stamens, however; but the second whorl has disappeared; while the pistil now[157] contains only one seed instead of three; though it still retains some trace of the original three cells in the fact that there are three sensitive surfaces, united together at their base into one stalk or style. Each such diminution in the number of seeds is always accompanied by an increase in the effectiveness of those which remain; the difference is just analogous to that between the myriad ill-provided eggs of the cod, whose young fry are for the most part snapped up as soon as hatched, and the two or three eggs of birds, which watch their brood with such tender care, or the single young of cows, horses, and elephants, which guard their calves or foals almost up to the age of full maturity. What the bird or the animal effects by constant feeding with worms or milk, the[158] plant effects by storing its seed with assorted food stuffs for the sprouting embryo.
In the more advanced or more degenerate sedges we get still further differentiation for the special function of wind-fertilisation. Take, as an example of these most developed types on this line of development, the common English group of carices (fig. 37). In these, the flowers have absolutely lost all trace of a perianth (that is to say, of the calyx and petals), for they do not possess even the six diminutive bristles which form the last relics of those organs in their allies, the scirpus group. Each flower is either male or female—that is to say, it consists of stamens or ovaries alone. The male flowers are represented by a single scale or bract, inclosing three stamens; and in some species even the stamens are reduced to a pair, so that all trace of the original trinary arrangement is absolutely lost. The female flowers are represented by a single ovary, inclosed in a sort of loose bag, which may perhaps be the final rudiment of a tubular bell-shaped corolla like that of the hyacinth. This ovary contains a single seed, but its shape is often triangular, and it has usually three stigmas or sensitive surfaces, thus dimly pointing back to the three distinct cells of its lily-like ancestors, and the three separate ovaries of its still earlier[159] alisma-like progenitors. In many species, however, even this last souvenir of the trinary type has been utterly obliterated, the ovary having only two stigmas, and assuming a flattened two-sided shape. In all the carices, the flowers are loosely arranged in compact spikes and spikelets, with their mobile stamens hanging out freely to the breeze, and their feathery stigmas prepared to catch the slightest grain of pollen which may happen to be wafted their way by any passing breath of air. The varieties in their arrangement, however, are almost as infinite among the different species as those of the grasses themselves; sometimes the male and female flowers are produced on separate plants; sometimes they grow in separate spikes on the same plant; sometimes the same spike has male flowers at the top and female at the bottom; sometimes the various flowers are mixed up with one another at top and bottom in a regular hotch-potch of higgledy-piggledy confusion. But all the sedges alike are very grass-like in their aspect, with thin blades by way of leaves, and blossoms on tall heads as in the grasses. In fact, the two families are never accurately distinguished by any except technical botanists; to the ordinary observer, they are all grasses together, without petty distinctions of genus and species. Like the grasses, too, the sedges[160] are mostly plants of the open wind-swept plains or marshy levels, where the facilities for wind-fertilisation are greatest and most constantly present.[6]
And now, from this illustrative digression, let us hark back again to the junction point of the rushes, whence alike the sedges and the grasses appear to diverge. In order to understand the nature of the steps by which the cereals have been developed from rush-like ancestors, it will be necessary to look shortly at the actual composition of the flower in grasses, which is the only part of their organism differing appreciably from the ordinary lily type. The blossoms of grasses, in their simplest form, consist of several little green florets, arranged in small clusters, known as spikelets, along a single common axis. Of this arrangement, the head of wheat itself offers a familiar and excellent example. If we pull to pieces one of the spikelets composing such a head, we find it to consist of four or five distinct florets. Omitting special features and unnecessary details, we may say that each floret is made up of two chaffy scales (e, d), known as pales, and representing the calyx, together with a pair[161] of small white petals (c) known as lodicules, three stamens (b), and an ovary with two feathery styles (a). Moreover, the two pales or calyx-pieces are not similar and symmetrical, for the outer one (e) is simple and convex, while the inner one (d) is apparently double, being made up of two pieces rolled into one, and still possessing two green midribs, which show distinctly like ribs on its flat outer surface. Here, it will immediately be apparent, the traces of the original trinary arrangement are very slight indeed.
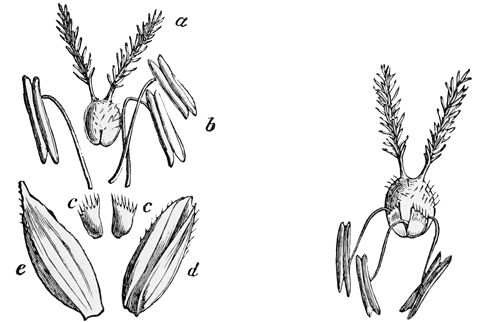
| Fig. 38. | Fig. 39. |
| Details of flower of Wheat. | Flower of Wheat (glumes removed). |
But when we come to inquire into the rationale and genesis of these curiously one-sided flowers, it is not difficult to see that they have been ultimately[162] derived from trinary blossoms of the rush-like type. The first and most marked divergence from that type, for which the analogy of the sedges has already prepared us, is the reduction of the ovary to a single one-seeded cell, whose ripe fruity form is known as a grain. At one time, we may feel pretty sure, there must have existed a group of nascent grasses, which only differed from the wood-rush genus in having a single-celled ovary instead of a three-celled pistil with one seed in each cell; and even the ovary of this primitive grass must have retained one mark of its trinary origin in its possession of three styles to its one grain, thus pointing back (as most sedges still do) to its earlier rush-like origin. That hypothetical form must have had three sepals, three petals, six stamens, and one three-styled ovary. But the peculiar shape of modern grass-flowers is clearly due to their very spiky arrangement along the edge of the axis. In the wood-rushes and the sedges, we see some approach to this condition; but in the grasses, the crowding is far more marked, and the one-sidedness has accordingly become far more conspicuous. Suppose we begin to crowd a number of wind-fertilised lily-like flowers along an axis in this manner, taking care that the stamens and the sensitive feathery styles are always turned outward to catch the breeze (for otherwise they[163] will die out at once), what sort of result shall we finally get?
In the first place, the calyx, consisting of three pieces, will stand towards the crowded stem or axis in such a fashion that one piece will be free and exterior, while two pieces will be interior and next the stem, thus—
Now, the effect of constant crushing in this direction will be that the two inner calyx-pieces will be slowly dwarfed, and will tend to coalesce with one another; and this is what has actually happened with the inner pale of wheat and of other grasses, though the midribs of the two originally separate pieces still show on the compound pale, like dark green lines down its centre. Thus, in the fully developed grasses, in place of a trinary calyx, we get two chaffy scales or pales, the outer one representing a single sepal, and the inner one, which has been dwarfed by pressure against the stem, representing two sepals rolled into one, with two midribs still remaining as evidence of their original distinctness.
Next, in the case of the petals, which alternate with the sepals of the calyx, the relation to the stem is exactly reversed; for we have here two petals free[164] and exterior, with one interior petal crowded closely against the axis, thus—
Here, then, the two external petals will be saved, exactly as the one external sepal was saved in the case of the calyx; and these two petals are represented by the very small white lodicules under the outer pale in our existing wheats and grasses. On the other hand, the inner petal, jammed in between the grain and the inner pale (with the stem at its back), has been utterly crushed out of existence, partly because of its very small size, partly because of its functional uselessness, and partly because it had no other part with which to coalesce, and so to save itself as the inner sepals had managed to do. Moreover, it must be remembered that the sepals do still perform a useful service in protecting the young flower before it opens, and in keeping out noxious insects during the kerning or swelling of the grain; whereas the lodicules or rudimentary petals are now apparently quite functionless; and so we may congratulate ourselves that they are there at all, to preserve for us the true ground-plan of the floral architecture in grasses. Indeed, they have not survived by any means in all grasses: among the smaller and more [165] degraded kinds they are often wholly wanting, having been quite crushed out between the calyx and the grain. It is only the larger and more primitive types that still exhibit them in any great perfection. On the other hand, one group of very large exotic grasses, the bamboos, has three regular petals, thus clearly showing the descent of the family as a whole from rush-like ancestors, and also obviously suggesting that the obsolescence of the inner petal in the other grasses is due to their small size and their closely packed minute flowers.
Among the stamens, one-sidedness has not notably established itself, for in wind-fertilised plants they must necessarily hang out freely to the breeze, and therefore they do not get much crowded between the other parts. A few grasses still even retain their double row of stamens, having six to each floret; but most of them have only one whorl of three. In some of the lower and more degraded forms, however, even the stamens have lost their trinary order, and only two now survive. This is the case in our own very degenerate little sweet-vernal-grass, the plant which imparts its delicious fragrance to new-mown hay. But in the cereals and in most other large species the three stamens still remain in undiminished effectiveness to the present day.
Finally we come to the most important part of all, the ovary. This part, alternating with the stamens, has the same arrangement of styles relatively to the axis as in the case of the petals; and it has undergone precisely the same sort of abortive distortion. The two outer styles, hanging freely out of the calyx, have been preserved like the two outer lodicules; but the inner one, pressed between the grain and the inner pale (with the stem behind it) has been simply crushed out of existence, like its neighbour the inner lodicule.
Thus the final result is that the whole inner portion of the flower (except as regards stamens) has been distorted or rendered abortive by close pressure against the stem (due to the crowding of the florets in the spiky form), while the whole outer portion remains normal and fully developed. We have an outer pale representing a single normal sepal, and an inner pale representing two dwarfed and united sepals; we have two normal outer lodicules or petals, and a blank where the inner petal ought to be; we have three stamens, symmetrically arranged, among the faithless faithful only found; and we have finally two normal outer styles, with a blank in place of the absent inner style. The accompanying diagram, compared with that of the[167] primitive monocotyledon (fig. 32), will make this perfectly clear.
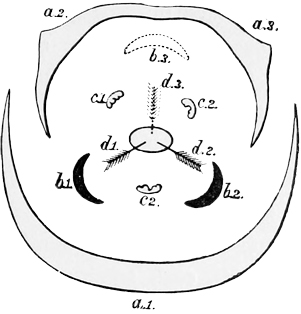
Fig. 40.—Diagram of Wheat flower.
Here, a1 represents the outer pale or normal sepal, while a2 and a3 represent the inner pale composed of the two united sepals. Again, b1 and b2 stand for the two lodicules or surviving petals, while b3 marks the place of the lost petal, now found in the bamboos alone. The stamens are lettered c1, c2, and c3. The two existing styles are shown by d1 and d2, while d3 marks the abortive inner style, now not even present in a rudimentary condition. It will be observed at once that all the outer side is normal, and all the inner side more or less abortive through pressure against the axis.
Thus it will be seen that the line of links which[168] connects the grasses and cereals with the lilies is absolutely unbroken, and that it consists throughout of one continuous course of degradation. At the same time, by this one-sided and spiky arrangement, the grasses secured for themselves an exceptional advantage in the struggle for existence. No other race of small wind-fertilised plants could compete with them for the possession of the open wind-swept plains; and over all these they spread far and wide, rapidly differentiating themselves into a vast number of divergent genera and species, each adaptively specialised for some peculiar habitat, soil, or climate. At the present time, the grasses number their kinds by thousands; they extend over the whole world from the poles to the equator; and they form the general sward or carpet of greenery over by far the larger portion of the terrestrial globe. Even in Britain alone, with our poor little insular flora, a mere fragment of that belonging to the petty European continent, we number no less than forty-two genera of grasses, distributed into more than one hundred species. In fact, what may fairly be called degradation from one point of view may fairly be called adaptation from another. The organisation of the grasses is certainly lower than that of the lilies, but it fits them better for that station of life to which it has pleased nature to assign them.
The various kinds of grasses differ very little from one another in general plan; the flower in almost all is constructed strictly on the lines above mentioned; and the leaves in almost all are just the same soft pensile blades, making them into the proper green sward for open, unwooded, wind-swept plains. But like almost all other very dominant families, they have split up into an immense number of kinds, distinguished from one another by minute differences in the arrangement of the florets and the spikelets; and these kinds have again subdivided into more and more minutely different genera and species. One great group, with panicles of a loose character, and very degraded spikelets, has given origin to many southern grasses, from some of which the cultivated millets are derived. Another great group, with usually more spiky inflorescence, has given origin to most of our northern grasses, from some of which the common cereals are derived. This second group has again split up into several others, of which the important one for our present purpose is that of the Hordeineæ, or barley-worts. From one of the numerous genera into which the primitive Hordeineæ have once more split up, our cultivated barleys take their rise; from another, which here demands further attention, we get our cultivated wheats.
The nearest form to true wheat now found wild in the British Isles is the creeping couch-grass, a perennial closely agreeing in all essential particulars of structure with our cultivated annual wheats. But in the south European region we find in abundance a large series of common wild annual grasses, forming the genus Ægilops of technical botany, and exactly resembling true wheat in every point except the size of the grain. One species of this genus, Ægilops ovata, a small, hard, wiry annual, is now pretty generally recognised among botanists as the parent of our cultivated corn. There was a good reason, indeed, why primitive man, when he first began to select and rudely till a few seeds for his own use, should have specially affected the grass tribe. No other family of plants has seeds richer in starches and glutens, as indeed might naturally be expected from the extreme diminution in the number of seeds to each flower. On the other hand, the flowers on each plant are peculiarly numerous; so that we get the combined advantages of many seeds, and rich seeds, so seldom to be found elsewhere except among the pulse family. The experiments conducted by the Agricultural Society in their College Garden at Cirencester have also shown that careful selection will produce large and rich seeds from Ægilops ovata, considerably resembling true wheat, after only a few years’ cultivation
Primitive man, of course, did not proceed nearly so fast as that. Of the very earliest attempts at cultivation of Ægilops, all traces are now lost, but we can gather that its tillage must have continued in some unknown western Asiatic region for some time before the neolithic period; for in that period we find a rude early form of wheat already considerably developed among the scanty relics of the Swiss lake dwellings. The other cultivated plants by which it is there accompanied, and the nature of the garden weeds which had followed in its wake, point back to Central or Western Asia as the land in which its tillage had first begun. From that region the Swiss lake dwellers brought it with them to their new home among the Alpine valleys. It differed much already from the wild Ægilops in size and stature; but at the same time it was far from having attained the stately dimensions of our modern corn. The ears found in the lake dwellings are shorter and narrower than our own; the spikelets stand out more horizontally, and the grains are hardly more than half the size of their modern descendants. The same thing is true in analogous ways with all the cultivated fruits or seeds of the stone age: they are invariably much smaller and poorer than their representatives in existing fields or gardens. From that time to this the[172] process of selection and amelioration has been constant and unbroken, until in our own day the descendants of these little degraded lilies, readapted to new functions under a fresh régime, have come to cover almost all the cultivable plains in all civilised countries, and supply by far the largest part of man’s food in Europe, Asia, America, and Australia.
The path up from the Llyn to the crest of Mynydd Mawr leads for some distance along the mossy, boulder-strewn course of a mountain torrent, which takes its rise in a fairy spring close below the actual summit of the craggy peak. It is a stiff pull for fair-weather pedestrians, this almost untrodden tourist trackway, with here and there a hand-and-knee clamber over great glacier-marked bosses of solid granite; but the exquisite glimpses we get at every fresh spur over the bare shoulders of Moel Siabod and into the cleft valley of the upper Conway more than compensate for the rough stony walking and the obvious damage to one’s nether integuments. Very few casual beaten-road visitors ever find out these lonely footpaths up the less-frequented mountains; the mass takes its circular tour round the regulation road by Llanberis, Beddgelert, and Capel Curig, leaving Mynydd Mawr and its neighbouring Carnedds[174] out in the cool shade of popular neglect. So much the better for those wandering naturalists who love to ramble among unhackneyed scenes, and to spy out wild nature in all her native loveliness, an Artemis who only bares her beauty among the deepest and most secret recesses of glade or woodland.

Fig. 41.—Lloydia serotina
(Mountain Tulip).
Here by the bank of the tiny torrent, where I shall stop and rest on a smooth stretch of naked rock for a few idle minutes, there is beauty enough in all conscience to charm the spellbound eyes of any intrusive Actæon. The moist fissures of the water-worn granite are richly clad with filmy fronds of alpine ferns; the drier crevices among the tumbled rocks are tufted with the black stems and graceful foliage of the maidenhair spleenwort; and the scanty alluvial mould on the slopes beyond is carpeted by lithe creeping sprays of beautiful branching club-moss. All[175] around me, a wealth of luxuriant mountain vegetation covers the peaty soil of the hollows, or the shallow granitic clay washed down into the crannies from the weathering crags above. There are insect-eating sundews, with their clammy red-haired leaves inclosing the half-digested bodies of a dozen tiny flies, whose attention they have falsely attracted with their delusive show of pretended honey. There are equally deceptive butterworts, with tall scapes of bright blue blossoms, and with pale yellowish-green foliage curled tightly round their mouldering victims in a deadly embrace. There are Alpine saxifrages, unfolding their pretty pinky-white flowers to the eager advances of the fertilising bees. And here amongst them all, in a sheltered nook of the inclosing granite débris, is the great prize of the day, the wee slender mountain tulip, in search of which I have come out this breezy morning, and whose actual home on the side of Mynydd I hardly expected to light upon so easily or so quickly in the upward march.
Of course I was told beforehand exactly where to look for it by the torrent’s brink; for our botanists have long ago so thoroughly overhauled every inch of England, Scotland, Wales, and Ireland, in search of specimens, that every individual station for every rare British plant is perfectly well known to them,[176] and printed in minute detail in half a dozen British floras. But I feared that here our little mountain tulip might be quite extinct already, exterminated by the too pressing attention of its numerous dilettante admirers; for as soon as your average collector finds a last lingering relic of some moribund British race on down or moorland, his first notion is to complete its destruction by rooting up the one remaining individual as a unique specimen, to become a permanent record of his luck and skill in the brown paper treasuries of his own herbarium. We, however, are naturalists of another kidney, I trust: we will observe and examine our little treasure carefully on the spot, but we will not pull it up ruthlessly, bulb and all, or press its pretty blossoms under a dead weight of books and drying paper, in order to preserve its miserable mummy in the wretched cemetery of a hortus siccus. Long may it flourish on its native hill-side, and may no scientific hand ever grub it up as the cruel trophy of a specimen-slaughtering raid! Indeed, to be perfectly frank, like the canny Scot who was ‘no thot sure of Jocky,’ I have not trusted even my readers themselves with the exact secret of my tulip’s whereabouts. I will confess that I have invented the name of Mynydd Mawr on purpose to deceive, and I have led up to the summit by a round[177]about path through the glen of Conway in order to prevent any future intruder from retracing his steps without me, and annexing for his own private aggrandisement the pretty flower whose life I have so chivalrously and humanely spared. When we come to learn the history of its race, I feel sure every one will sympathise in the sentiment which makes me wish to preserve this solitary colony of Alpine flowers as long as possible from the desecrating hands of the abandoned plant-collector.
First, let us look exactly what manner of lily it really is, and then we will go on to unravel together the clues and tokens of its romantic history. See, it is a little simple grass-like plant, sending forth from its buried bulb two or three very slender blades by way of leaves; and from their midst springs a graceful bending stem, surmounted by a single star-shaped white blossom. At least, it looks white at first sight, though when you come to examine it more closely you can observe three red lines running down the face of each petal, and converging on a small bright golden spot at their base. Those lines are in fact honey-guides for the mountain insects, pointing them the shortest road to the sweets stored up in the nectaries, and so saving them any extra trouble in looking about for their morning’s meal. On the other[178] hand, the insects repay the flower for its honey by carrying pollen from blossom to blossom, and so enabling the plant to set its seed. Of course, unless the young capsule in the centre of each blossom is thus fertilised by pollen from one of its neighbours, it never ripens into a seed-bearing fruit at all; and, indeed, in the economy of the plant itself, the sole object of the blossom, with its bright petals, its store of honey, and its faint perfume (almost imperceptible to any save very delicate senses), is simply to induce the bee or the butterfly thus to convey the fertilising powder from one head of flowers to another of the same sort.
Our little plant has of course a botanical name of the usual clumsy kind; but in this particular instance there is a certain rough fitness in its application, for being a Welsh lily by nature it is duly known by a Latinised Welsh name as Lloydia. Now, I am not going this morning to inquire fully into the whole past history of the original family from which it springs—that would be too long a subject for an off-hand lecture as I sit here basking on the bare granite slope; I propose only entering in any detail into the last chapter of its chequered career, and asking how it has managed to keep its foothold for so many ages in this one spot and on a few neighbouring Snow[179]donian summits. But before we go into that final question we must just begin, by way of preparatory exercise, with a very brief account of its earlier origin. Lloydia serotina, then, to give it the full benefit of its Latinised name, is a mountain plant of northern and Arctic Europe, as well as of the chillier portions of Siberia and British North America. Further south, it is found only in the colder upland shoulders of the Alps, the Caucasus, and the Altai range, as well as in a few other great snowy mountain systems; but in Britain it occurs nowhere except on one or two of the higher mountains here in North Wales. By origin, it is a very early and simple offshoot of the great lily tribe; one of the most primitive lilies, indeed, now existing on the face of the earth. Like all others of that vast family, it has six petals and six stamens or pollen-bearing sacs; but it still retains a very early form of lily flower in its open star-shaped blossom as well as in one or two other smaller peculiarities. The cultivated tulips of our gardens, varieties of a wild Levantine species, are all descended from a somewhat similar form; but with them the course of development has gone much further; the petals have grown far larger and more conspicuous, in order to allure the eyes of bigger southern insects, and the general form of the flower has become bell-shaped[180] instead of star-shaped, in order to ensure more safe and certain fertilisation by these winged allies; for in a tubular blossom the pollen is much more likely to be brushed off from the insect’s head on to the proper portion of the unripe capsule than in an open spreading flower like our Lloydia here. Hence we may fairly say that Lloydia represents an early ancestral form from which the modern and more southerly tulips are nature’s enlarged and improved varieties.
But how did these pretty little white lilies get here, and why do they still remain here in their early simple form, while their southern sisters elsewhere have been slowly modified into brilliant yellow bell-shaped tulips? Thereby hangs a most curious and delightful tale. For I have very little doubt that the ancestors of our pretty lilies here have been growing uninterruptedly in the present spot for many thousands of years, and that during all that time they have gone on reproducing themselves by seed from time to time, without once having crossed their stock with any of their congeners in the Arctic regions or in the great snow-clad ranges of central Europe. Indeed, I very much doubt whether they have ever even intermarried with their neighbours on the other Snowdonian summits, for I think I shall be able to show good reasons for believing that each of these[181] little isolated colonies has lived on for ages all by itself on each of their three scattered peaks in the North Welsh district.
It is a curious fact, certainly, that one should find a single species of Arctic flower reappearing at such long distances in such isolated spots under closely similar circumstances. If we go to the great snow-clad stretches of land which extend around the Arctic Circle in Europe, Asia, and America, we shall everywhere find our little lily growing in abundance close up to the line of perpetual snow, though its diverse habitats are there divided by wide expanses of open sea. If, again, we cross the whole of the German plains, we shall see no Lloydias in the intervening tract; but when we reach the Alps and the Pyrenees, we shall a second time come upon other isolated colonies of the self-same flower. Once more, we may turn eastward, and we shall meet with it, after a long march, among the Carpathians and the Caucasus; or we may turn westward, and then we shall light upon it again on the craggy sides of a few solitary Welsh mountains. How does it come that in every cold tract we find the self-same species recurring again and again wherever the circumstances are fitted for its growth? and how have its seeds or bulbs been conveyed across such wide stretches of inter[182]vening sea or valley to so many distinct and separate chilly regions?
One obvious answer might be, that under similar conditions a like flower had everywhere been developed from some common plant of lowland or temperate districts. But in reality such absolute similarity of independent development never actually occurs in nature, for the various Lloydias are not merely rather like one another, but are actually one and the same species, as like each other (to quote our old Welsh friend Fluellin) ‘as my fingers is to my fingers.’ Now, naturalists know that such absolute identity of structure can only arise through unbroken descent from a common origin; wherever two species are separately descended from unlike ancestors, however close their analogies may be, they are always at once marked off from one another by some very obvious points of structural dissimilarity. Nor can we suppose that the seeds of the Lloydia have been transported from one place to another by mere accident, clinging to the legs of Arctic birds, or carried unwittingly on the muddy heels of globe-trotting tourists. Such accidents do indeed occasionally occur, and they account for the very fragmentary manner in which remote Oceanic islands like the Azores or St. Helena are peopled by waifs and[183] strays from the fauna and flora of all neighbouring continents. But as we have already seen in the converse case of the hairy spurge, it would be incredible that such an accident should have occurred over and over again in a hundred separate cases, so that every suitable place in the whole northern hemisphere should separately, by mere luck, have received a distinct colony of appropriate cold-climate plants. Incredible, I should say, even if the instance of the Lloydia stood alone without any analogues; but in fact, as I shall try to point out by-and-by, it is only one instance out of a thousand that might be quoted; for every Arctic land and every snow-clad Alpine peak is covered close up to the limit of vegetation with dozens or hundreds of similar plants, insects, and animals, which are nowhere found in all the intervening temperate or lowland regions. Clearly all these coincidences cannot be due to mere accident; we must seek for their reason in some single and common fact.
See this great rounded block of smooth granite on whose solid shoulders I am now sitting; how wonderfully grooved and polished it is, with long, deep, rounded furrows running lengthwise across its face in the same direction as the general dip of the Conway valley. What can have made those curious[184] parallel channels on its naked surface, I wonder? Any one who has ever looked closely at the rocks about the foot of a glacier in Switzerland will recognise at once what was the agency at work on the granite slopes of Mynydd Mawr. Those are most undoubtedly ice-marks, caused by the long, slow, grinding action of the superincumbent glaciers. For of course everybody knows nowadays that there was once a time when great glacial sheets spread over the combes and glens of Snowdonia, as they spread to-day over the nants of Chamounix and the buried basin of the Mer de Glace.
Dr. Croll’s calculations have shown that the astronomical conjunction necessary for the production of such a state of things must have occurred some two hundred thousand years since; and from that date down to eighty thousand years ago our planet kept presenting alternately either pole to the sun during long cycles of 10,500 years each; so that, first, the northern hemisphere enjoyed a long summer, while the southern was enveloped for a vast distance from the Antarctic Circle in a single covering sheet of ice; and then again the southern hemisphere had its lengthened spell of tropical weather, while the north was turned into one enormous Greenland down as far as the British Isles. Eighty thousand years ago,[185] or thereabouts, this condition of things began to change; the climate of the north became more genial; and ever since that date our sober planet has oscillated within gentler limits, producing only such alternate results of annual summer and winter as those with which we ourselves are now familiar.
When the glaciation was at its worst in the northern hemisphere, almost the entire surface of the European continent, from Scandinavia and Lapland, to England, Belgium, and central Germany, lay buried beneath one unbroken sheet of permanent ice. But when the conditions were a little less severe, local glaciers radiated from the chief mountain bosses of Scotland, Wales, and the Isle of Man, and ground these deep grooves and scratches on the worn surface of the denuded rock. At length the climate began to mend slightly; and then the plants and animals of the Arctic zone spread uninterruptedly over the whole of northern Europe, from the limit of pack-ice to at least the southern slope of the Alps on the Italian side. Remains of these glacial animals—Arctic lemmings, musk sheep, white hares, reindeers, Alpine marmots, and snowy owls—are still found among the bone-caves and river drift of the interglacial ages in various parts of Europe, from Scandinavia to the Tuscan grottoes. At the same time we may be pretty[186] sure that high Arctic or Alpine plants, adapted to a chilly climate, like the saxifrages, the sibbaldia, the crowberry, and the Swiss veronica, spread over all the plains and valleys of Great Britain and the neighbouring continent.
In those days, we saw good reason to believe when we were examining the stranded southern flora of Cornwall and Devonshire, England and Ireland were united to one another as well as to France and Holland by a broad belt of lowland occupying what is now the bed of the two channels and the German Ocean, so that the mammoth and the cave-bear could roam uninterruptedly from the Yorkshire hills to the rock-shelters of the Dordogne, and from the bogs of Connaught to the then ice-clad summits of the Hartz and the Jura. The dark hunters of the period, who framed the rough, chipped stone hatchets of the Abbeville drift and the beautiful flint arrowheads of the southern French caves, could in like manner range without let or hindrance from Kent’s Hole at Torquay to the Schwatka cavern in Moravia, and from the honeycombed cliffs of Yorkshire valleys to the limestone grottoes among the Alpine slopes. That distribution of land and water easily accounts for the dispersion of Arctic and snow-line plants or animals over all the snowy regions of northern Europe.
But as the cold began to subside, and as a warmer fauna and flora invaded the now milder plains and valleys of central Europe, the glacial types, being less adapted to the new conditions, began to retreat northward towards the Arctic Circle, or upward towards the chilly summits of the principal mountains. Slowly, age after age, the southern plants and animals overran all the lower portions of the continent, cutting the glacial fauna and flora in two, and established themselves as far as the outlying peninsula of Britain, which still continued to form an integral part of the European mainland. After most of the Germanic types had made good their foothold even in this distant region, however, and after the still more southern pæonies of the Steep Holme and the rock-cistus of Torquay had established themselves under the lee of the Cornish and Kerry mountains, on the submerged tract which then stretched out far to the west of the Scilly Islands, the land began to sink slowly toward sea-level; and at last an arm of the Atlantic encircled the whole of Ireland, and still later the waters of two long gulfs which now form the English Channel and the North Sea met together by bursting through the narrow barrier of chalk between Dover and Cape Blancnez. Thus Britain finally became an island group; and, being washed on three sides by the warm[188] current of the Gulf Stream, it acquired an unusually high and equable temperature for a district situated so far to the north and rising into so many chains of low mountains. But not all the plants and animals which inhabit the continent had had time to reach England, which has a comparatively poor fauna and flora; while still more failed to get to Ireland before the separation; and so, the Irish flora contains a larger proportion of Spanish and Portuguese types, while the mass of the English flora, especially in the eastern half of the island, is essentially Germanic.
Even after this change to more genial conditions, however, many of the Arctic plants, though now separated by wide stretches of sea or land from their nearest relatives elsewhere, managed to keep up a vigorous existence in the Scotch Highlands, the Welsh hills, and the greater summits of the Lake district. Some of them still cover vast tracts of country in the north; as, for example, the little green sibbaldia, a tufted Arctic trailer, whose herbage forms a chief element of the greensward in many parts of the Highlands; or the pretty eight-petalled dryas, which stars with its sweet white blossoms the limestone rocks of northern England and the Ulster hills. Among the more common of these isolated old glacial flowers in Britain are the Alpine meadow-rue, the[189] northern rock-cress, the Arctic whitlow-grass, the Alpine pearlwort, the Scottish asphodel, the mossy cyphel, the mountain lady’s mantle, the purple saxifrage, and the red bearberry. Altogether, we have still more than two hundred such Alpine or Arctic plants, stranded among our uplands or in the extreme north of Scotland, and probably separated for many thousand years from the main body of their kind in the Arctic Circle or the snowy mountains of central Europe.
Our pretty little Lloydia here is far rarer in Britain than these low mountain kinds; for it has died out utterly even in Scotland itself, and now survives nowhere with us except on these solitary Welsh summits. Such cases are frequent enough in Britain; for while the moderate mountainous or Arctic species still go on thriving among the straths and corries, the coldest kinds of all have often been pushed upward and ever upward by the advancing tide of southern flowers till they are left at last only on a few isolated mountain tops, where many of them are even now in course of slowly disappearing before the steady advance of the southern types. For example, there is a certain pretty kind of heath, confined to northern or Arctic hill-sides, which till lately lingered on in Britain only on the one mountain known as the Sow of Athole in Perthshire;[190] but of late years it has grown rarer and rarer with each succeeding summer, until it is now probably quite extinct. It is the natural tendency of all such small isolated colonies, whether of plants or animals, to die slowly out; for they cannot cross freely with any of their own kind outside the narrow limits of their own restricted community; and by constantly breeding in and in with one another they at last acquire such weak and feeble constitutions that they finally dwindle away imperceptibly for want of a healthy infusion of fresh external blood.
Fig. 42.—Lady’s Slipper (Cypripedium calceolius).
If I mention a few other like cases (as well as I can remember them on the spur of the moment, for I cannot pretend to give a complete ex-cathedrâ list here on the slopes of Mynydd Mawr) it will help to elucidate the origin and nature of this little colony of mountain tulips. There is a lovely orchid, the lady’s slipper, common in Siberia and Russia, almost up to the Arctic Circle, but now found with us only in one Yorkshire station, where, like the Perthshire heath, it is rapidly verging to complete local extinction. Again, among one family alone, the tufted saxifrage has now been driven to the summits of Ben Avers and Ben Nevis; the drooping saxifrage is extinct everywhere in Britain save on the cloudy top of Ben Lawers; the[191] brook saxifrage lingers on upon the same mountain, as well as on Ben Nevis and Lochnagar; and the Alpine saxifrage, though more frequent in little solitary groups in Scotland and the Lake district, has died out[192] of all Ireland save only on the bald head of Ben Bulben in Sligo. The Alpine sow-thistle, an Arctic and snowy weed, is now dying out with us on the tops of Lochnagar and the Clova mountains. The black bearberry yet haunts Ben Nevis and a few other Highland peaks. The Alpine butterwort has been driven even from the mountains in Scotland generally, but still drags on a secluded existence in a few very northern bogs of Caithness and Sutherland; in this respect it resembles the northern holy-grass, an Arctic plant, which Robert Dick, the self-taught botanist of Thurso, discovered among the high pastures near his native town. This same grass strangely reappears in New Zealand, whither it has doubtless been carried from Siberia by its seeds accidentally clinging to the feet of some belated bird; but then such a solitary case in itself shows how impossible is the explanation of the numerous Scotch and Welsh Arctic plants as due to mere chance; for while in north European mountains similar instances can be counted by hundreds, in New Zealand the coincidence is very rare and almost unparalleled.
The snowy gentian, to continue our list, turns up in a good many little Scotch colonies; but the Alpine lychnis, its companion on the mountain pastures of the Bernese Oberland, is only now known in Britain[193] on the summit of Little Kilrannoch, a Forfarshire mountain, and among the crags of Hobcartin Fell in Cumberland. The bog sandwort, everywhere a rare and dying species, has wholly disappeared from these islands except on the sides of the Widdybank Fell in Durham. Its ally the fringed sandwort loiters late on the limestone cliffs of Ben Bulben in Sligo, as well as on one solitary serpentine hill in the island of Unst among the chilly Shetlands. A tiny pea-flower, the Alpine astragalus, has been driven almost everywhere to the snow-line, but still survives in Scotland among the Clova and Braemar mountains. It is on a single spot in the same exposed Clova range, too, that the closely related yellow oxytrope still grows in diminishing numbers; while its ally the Ural oxytrope holds its own manfully over all the dry hills of the Highlands. I could add to these instances many more; but lunch is waiting to be eaten in the knapsack, and I am loth to tire the patience of my hearers with too long a list of barren names and bare wind-swept mountain summits.
Still, I love to think that the little colony of timid shrinking Lloydias stranded here on the granite slopes of Mynydd Mawr can push back its pedigree in such an unbroken line to so dim and distant a prehistoric past. Ever since the glaciers last cleared away from[194] this boss of smooth stone on whose broad back we are sitting, a tiny group of our pretty mountain tulips has continuously occupied age after age this self-same spot. Originally, no doubt, they covered the whole sides of the mountains and stretched down far into the plains and valleys; but gradually, as the world’s weather grew warmer, they were restricted, first, to the mountain tracts of Wales and Scotland, then to a small Snowdonian district, and finally, even within that shrunken realm, to two or three isolated peaks. Occasionally, I suppose, a seed from one of the three existing Welsh colonies may be carried by accident into the territory of the others; but it is in the highest degree improbable that the stock has ever been reinforced for the last fifty thousand years from any purely external body of its congeners in the higher Alps or in the Arctic regions. The dark small men of the neolithic age, the Aryan Celts of the bronze period, the conquering Roman from the south, the Englishman, the Scandinavian, the Norman, all have since come, and most of them have gone again; but the Lloydias still hold precarious possession of their solitary remaining strongholds. An analogy from the animal world will help to bring out the full strangeness of this extraordinary isolation. Mount Washington in New Hampshire is the highest peak among[195] the beautiful tumbled range of the White Mountains. On and near its summit a small community of butterflies belonging to an old Glacial and Arctic species still lingers over a very small area, where it has held its own for the eighty thousand years that have elapsed since the termination of the great ice age. The actual summit of the mountain rises to a height of 6,293 feet; and the butterflies do not range lower than the five thousand feet line—as though they were confined on Snowdon to a district between the Ordnance cairn and the level of the little slumbering tarn of Glasllyn. Again, from Mount Washington to Long’s Peak in Colorado, the distance amounts to 1,800 miles; while from the White Mountains to Hopedale in Labrador, where the same butterflies first reappear, makes a bee-line of fully a thousand miles. In the intervening districts there are no insects of the same species. Hence we must conclude that the few butterflies left behind by the retreating main-guard of their race on that one New Hampshire peak have gone on for thousands and thousands of years, producing eggs and growing from caterpillars into full-fledged insects, without once effecting a cross with the remainder of their congeners among the snows of the Rocky Mountains or in the chilly plains of sub-Arctic America. So far as they themselves know, they are the only[196] representatives of their kind now remaining on the whole earth, left behind like the ark on Ararat amid the helpless ruins of an antediluvian world. Well, what these Mount Washington butterflies are among insects, that are our pretty wild tulips here among English flowers. They remain to us as isolated relics of an order that has long passed away; and they help us to rebuild with fuller certainty the strange half-undeciphered history of the years that were dead and gone long before written books had yet begun to be.
Fig. 43.—Common Cinquefoil.
Although the roses, like many other highly respectable modern families, cannot claim for themselves any remarkable antiquity—their tribe is only known, with certainty, to date back some three or four millions of years, to the tertiary period of geology—they have yet in many respects one of the most interesting and[198] instructive histories among all the annals of English plants. In a comparatively short space of time they have managed to assume the most varied forms; and their numerous transformations are well attested for us by the great diversity of their existing representatives. Some of them have produced extremely beautiful and showy flowers, as is the case with the cultivated roses of our gardens, as well as with the dog-roses, the sweet-briars, the may, the blackthorn, and the meadow-sweet of our hedges, our copses, and our open fields. Others have developed edible fruits, like the pear, the apple, the apricot, the peach, the nectarine, the cherry, the strawberry, the raspberry, and the plum; while yet others again, which are less serviceable to lordly man, supply the woodland birds or even the village children with blackberries, dewberries, cloudberries, hips, haws, sloes, crab-apples, and rowanberries. Moreover, the various members of the rose family exhibit almost every variety of size and habit, from the creeping silver-weed which covers our roadsides or the tiny alchemilla which peeps out from the crannies of our walls, through the herb-like meadow-sweet, the scrambling briars, the shrubby hawthorn, and the bushy bird-cherry, to the taller and more arborescent forms of the apple-tree, the pear-tree, and the mountain ash. And since modern science teaches[199] us that all these very divergent plants are ultimately descended from a single common ancestor—the primæval progenitor of the entire rose tribe—whence they have gradually branched off in various directions, owing to separately slight modifications of structure and habit, it is clear that the history of the roses must really be one of great interest and significance from the new standpoint of evolution. I propose, therefore, here to examine the origin and development of the existing English roses, with as little technical detail as possible; and I shall refer for the most part only to those common and familiar forms which, like the apple, the strawberry, or the cabbage rose, are already presumably old acquaintances of all my readers.
The method of our inquiry must be a strictly genealogical one. For example, if we ask at the present day whence came our own eatable garden plums, competent botanists will tell us that they are a highly cultivated and carefully selected variety of the common sloe or blackthorn. It is true, the sloe is a small, sour, and almost uneatable fruit, the bush on which it grows is short and trunkless, and its branches are thickly covered with very sharp stout thorns; whereas the cultivated plum is borne upon a shapely spreading tree, with no thorns, and a well[200]-marked trunk, while the fruit itself is much larger, sweeter, and more brightly coloured than the ancestral sloe. But these changes have easily been produced by long tillage and constant selection of the best fruiters through many ages of human agriculture. So, again, if we ask what is the origin of our pretty old-fashioned Scotch roses, the botanists will tell us in like manner that they are double varieties of the wild burnet-rose which grows beside the long tidal lochs of the Scotch Highlands, or clambers over the heathy cliffs of Cumberland and Yorkshire. The wild form of the burnet-rose has only five simple petals, like our own common sweet-briar; but all wild flowers when carefully planted in a rich soil show a tendency to double their petals; and, by selecting for many generations those burnet-roses which showed this doubling tendency in the highest degree, our florists have at last succeeded in producing the pretty Scotch roses which may still be found (thank Heaven!) in many quiet cottage gardens, though ousted from fashionable society by the Marshal Niels and Gloires de Dijon of modern scientific horticulturists.
Now, if we push our inquiry a step further back, we shall find that this which is true of cultivated plants in their descent from wild parent stocks, is true also of the parent stocks themselves in their[201] descent from an earlier common ancestor. Each of them has been produced by the selective action of nature, which has favoured certain individuals in the struggle for existence, at the expense of others, and has thus finally resulted in the establishment of new species, having peculiar points of advantage of their own, now wholly distinct from the original species whose descendants they are. Looked at in this manner, every family of plants or animals becomes a sort of puzzle for our ingenuity, as we can to some extent reconstruct the family genealogy by noting in what points the various members resemble one another, and in what points they differ among themselves. To discover the relationship of the various English members of the rose tribe to each other—their varying degrees of cousinship or of remoter community of descent—is the object which we set before ourselves in the present paper.
Perhaps the simplest and earliest type of the rose family now remaining in England is to be found in the little yellow potentillas which grow abundantly in ill-kept fields or by scrubby roadsides. The potentillas are less familiar to us than most others of the rose family, and therefore I am sorry that I am obliged to begin by introducing them first to my reader’s notice rather than some other and older[202] acquaintance, like the pear or the hawthorn. But as they form the most central typical specimen of the rose tribe which we now possess in England, it is almost necessary to start our description with them, just as in tracing a family pedigree we must set out from the earliest recognisable ancestor, even though he may be far less eminent and less well-known than many of his later descendants. For to a form very much like the potentillas all the rose family trace their descent. The two best known species of potentilla are the goose-weed or silver-weed, and the cinquefoil.[7] Both of them are low creeping herb-like weeds, with simple bright yellow blossoms about the size of a strawberry flower, having each five golden petals, and bearing a number of small dry brown seeds on a long green stalk. At first sight a casual observer would hardly take them for roses at all, but a closer view would show that they resemble in all essential particulars an old-fashioned single yellow rose in miniature. From some such small creeping plants as these all the roses are probably descended. Observe, I do not say that they are the direct offspring of the potentillas, but merely that they are the offspring of some very similar simple form. We ourselves do not derive our origin from the Icelanders; but the Ice[203]landers keep closer than any other existing people to that primitive Teutonic and Scandinavian stock from which we and all the other people of northwestern Europe are descended. Just so, the roses do not necessarily derive their origin from the potentillas, but the potentillas keep closer than any other existing rose to that primitive rosaceous stock from which all the other members of the family are descended.[8]
The strawberry is one of the more developed plants which has varied least from this early type represented by the cinquefoil and the silver-weed. There is, in fact, one common English potentilla, whose nature we have already considered, and which bears with village children the essentially correct and suggestive name of barren strawberry. This particular potentilla differs from most others of its class in having white petals instead of yellow ones, and in having three leaflets on each stalk instead of five or seven. When it is in flower only it is difficult at first sight to distinguish it from the strawberry blossom, though the petals are generally smaller, and the whole flower less widely opened. After blossoming, however, the[204] green bed or receptacle on which the little seeds[9] are seated does not swell out (as in the true strawberry) into a sweet, pulpy, red mass, but remains a mere dry stalk for the tiny bunch of small hard inedible nuts. The barren strawberry, indeed, as we saw in an earlier paper, is really an intermediate stage between the other potentillas and the true eatable strawberry; or, to put it more correctly, the eatable strawberry is a white-flowered potentilla which has acquired the habit of producing a sweet and bright-coloured fruit instead of a few small dry seeds. Since we have got to understand the rationale of this first and simplest transformation, we have now a clue by which we may interpret almost all the subsequent modifications of the rose family, and I must therefore be permitted here briefly to recapitulate the chief points we have already proved in this matter.
The true strawberry, we saw, resembles the barren strawberry in every particular except in its fruit. It is a mere slightly divergent variety of that particular species of potentilla, though the great importance of the variety from man’s practical point of view causes us to give it a separate name, and has even wrongly[205] induced botanists to place it in a separate genus all by itself. In reality, however, the peculiarity of the fruit is an extremely slight one, very easily brought about. In all other points—in its root, its leaf, its stem, its flower, nay, even its silky hairs—the strawberry all but exactly reproduces the white potentilla. It is nothing more than one of these potentillas with a slight diversity in the way it forms its fruit. To account, therefore, for the strawberry we had first to account for the white potentilla from which it springs.
The white potentilla, or barren strawberry, you will remember, is itself a slightly divergent form of the yellow potentillas, such as the cinquefoil. From these it differs in three chief particulars. In the first place, it does not creep, but stands erect; this is due to its mode of life on banks or in open woods, not among grass and meadows as is the case with the straggling cinquefoil. In the second place, it has three leaflets on each stalk instead of five, and this is a slight variation of a sort liable to turn up at any time in any plant, as the number of leaflets is very seldom quite constant. In the third place, it has white petals instead of yellow ones, and this is the most important difference of any. All flowers with bright and conspicuous petals we know are fertilised by insects, which visit them in search of honey or[206] pollen, and the use of the coloured petals is, in fact, to attract the insects and to induce them to fertilise the seeds. Now, yellow seems to have been the original colour of the petals in almost all (if not absolutely in all) families of flowers; and the greater number of potentillas are still yellow. But different flowers are visited and fertilised by different insects, and as some insects like one colour and some another, many blossoms have acquired white or pink or purple petals in the place of yellow ones, to suit the particular taste of their insect friends. In tracing the upward course of development in the roses, we shall see that they follow the ordinary law of progressive chromatic changes: the simpler types are yellow; the somewhat higher ones are white; the next pink; and the highest in this particular family are red; for no rose has yet attained to the final stage of all, which is blue. The colours of petals are always liable to vary, as we all see in our gardens, where florists can produce at will almost any shade or tint that they choose; and when wild flowers happen to vary in this way, they often get visited by some fresh kind of insect which fertilises their seeds better than the old ones did, and so in time they set up a new variety or a new species. Two of our English potentillas have thus acquired white flowers to suit their proper flies, while one[207] boggy species has developed purple petals to meet the æsthetic requirements of the marshland insects. No doubt the white blossoms of the barren strawberry are thus due to some original ‘sport’ or accidental variation, which has been perpetuated and become a fixed habit of the plant because it gave it a better and surer chance of setting its seeds, and so of handing down its peculiarities to future generations.
And now, how did we find the true strawberry had developed from the three-leaved white potentilla? Here the birds came in to play their part, as the bees and flies had done in producing the white blossom. Birds are largely dependent upon fruits and seeds for their livelihood, and so far as they are concerned it does not matter much to them which they eat. But from the point of view of the plant it matters a great deal. For if a bird eats and digests a seed, then the seed can never grow up to be a young plant; and it has so far utterly failed of its true purpose. If, however, the fruit has a hard indigestible seed inside it (or, in the case of the strawberry, outside it), the plant is all the better for the fact, since the seed will not be destroyed by the bird, but will merely be dispersed by it, and so aided in attaining its proper growth. Thus, if certain potentillas happened ever[208] to swell out their seed receptacle into a sweet pulpy mass, and if this mass happened to attract birds, the potentillas would gain an advantage by their new habit, and would therefore quickly develop into wild strawberries as we now get them. But the difference between the strawberry fruit and the potentilla fruit is to the last a very slight one. Both have a number of little dry seeds seated on a receptacle; only, in the strawberry the receptacle grows red and succulent, while in the potentilla it remains small and stalk-like. The red colour and sweet juice of the strawberry serve to attract the birds which aid in dispersing the seed, just as the white or yellow petals and the sweet honey of the potentilla blossoms serve to attract the insects which aid in fertilising the flowers. In this way all nature is one continual round of interaction and mutual dependence between the animal and vegetable worlds.
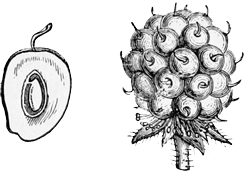
Fig. 44.—Fruit of Bramble.
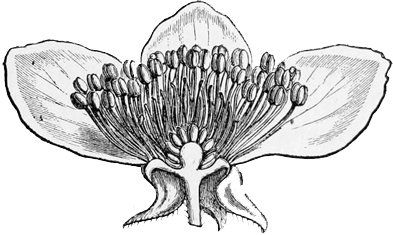
Fig. 45.—Flower of Bramble.
The potentillas and the strawberry plant are all of them mere low creeping or skulking herbs, without woody stems or other permanent branches. But when we get to the development of the brambles or blackberry bushes, we arrive at a higher and more respectable division of the rose family. There are two or three intermediate forms, such as water-avens and herb-bennet—tall, branching, weedy-looking[209] roadside plants—which help us to bridge over the gulf from the one type to the other. Indeed, even the strawberry and the cinquefoil have a short perennial, almost woody stock, close to the ground, from which the annual branches spring; and in some other English weeds of the rose family the branches themselves are much stiffer and woodier than in these creeping plants. But in the brambles, the trunk and boughs have become really woody, by the deposit of[210] hard material in the cells which make up their substance. Still, even the brambles are yet at heart mere creepers like the cinquefoil. They do not grow erect and upright on their own stems: they trail and skulk and twine in and out among other and taller bushes than themselves. The leaves remain very much of the cinquefoil type; and altogether there is a good deal of the potentilla left in the brambles even now.
However, these woody climbers have certainly some fresh and more developed peculiarities of their own. They are all prickly shrubs, and the origin of their prickles is sufficiently simple. Even the potentillas have usually hairs on their stems; and these hairs serve to prevent the ants and other honey-thieving insects from running up the stalks and stealing the nectar intended for fertilising bees and butterflies. Similar hairs in the goose-grass grew, you will recollect, into the little clinging hooks of the stems and midribs. But in the brambles, hairs of the same sort have grown still thicker and stouter, side by side with the general growth in woodiness of the whole plant; so that they have at last developed into short thorns, which serve to protect the leaves and stem from herbivorous animals. As a rule, the bushes and weeds which grow in waste places are very apt to be pro[211]tected in some such fashion, as we see in the case of gorse, nettles, blackthorn, holly, thistles, and other plants; but the particular nature of the protection varies much from plant to plant. In the brambles it takes the form of stiff prickly hairs; in the nettles, of stinging hairs; in the gorse, of pointed leaves; and in the thorn-bushes of short, sharp, barren branches.
Another peculiarity of the bramble group is their larger white flowers and their curious granulated fruit. The flowers, of course, are larger and whiter in order to secure the visits of their proper fertilising insects; the fruits are sweet and coloured in order to attract the hedgerow birds. Observe, too, that the flowers being higher in type than those of the strawberry, are often tinged with pink: here we get the first upward step in the direction of the true roses. The nature of the fruit in the raspberry, the blackberry, and the dewberry, again, is quite different from that of the strawberry. Here, instead of the receptacle swelling out and growing red and juicy, it is the outer coat of the separate little seeds themselves that forms the eatable part; while the receptacle remains white and inedible, being the ‘hull’ or stem which we pick out from the hollow thimble-like fruit in the raspberry. Each little nut, which in the strawberry was quite hard and brown, is here covered with a juicy black or[212] red pulp, inside which lies the stony real seed; so that a blackberry looks like a whole collection of tiny separate fruits, run together into a single head. Moreover, there are other minor differences in the berries themselves, even within the bramble group; for while the raspberry and cloudberry are red, to suit one set of birds, the blackberry and dewberry are bluish black, to suit another set; and while the little grains hold together as a cup in the raspberry, but separate from the hull, they cling to the hull in the other kinds. Nevertheless, in leaves, flower, and fruit there is a very close fundamental agreement among all the bramble kind and the potentillas. Thus we may say that the brambles form a small minor branch of the rose family, which has first acquired a woody habit and a succulent fruit, and has then split up once more into several smaller but closely allied groups, such as the blackberries, the raspberries, the dewberries, and the stony brambles.
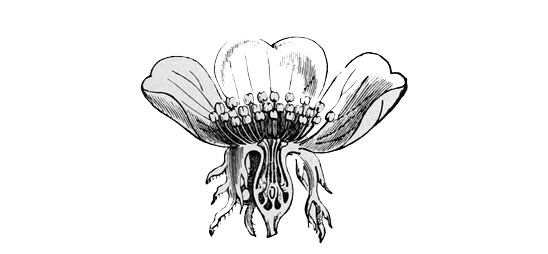
Fig. 46.—Vertical section of a Dog-rose.
The true roses, represented in England by the dog-rose and sweet-briar, show us a somewhat different development from the original type. They, too, have grown into tall bushes, less scrambling and more erect than the brambles. They have leaves of somewhat the same sort, and prickles which are similarly produced by the hardening of sharp hairs upon the[213] stem. But their flowers and fruit are slightly more specialised—more altered, that is to say, for a particular purpose from the primitive plan. In the first place, the flowers, though still the same in general arrangement, with five petals and many stamens and carpels (or fruit-pieces), have varied a good deal in detail. The petals are here much larger, and they have advanced to the stage of a brilliant pink; while the blossoms are also sweet-scented. These peculiarities of course serve to attract the bees and other large fertilising insects, which thus carry pollen from head to head, and aid in setting the seeds much more securely than the little pilfering flies. Moreover, in all the roses, the outer green cup which covers the blossom in the bud has grown up around the little seeds or fruit-pieces, so that instead of a ball turned outward, as in the strawberry and raspberry, you get,[214] as it were, a bottle turned inward, with the seeds on the inner side. After flowering, as the fruit ripens, this outer cup grows round and red, forming the hip or fruit-case, inside which are to be found the separate little hairy seeds. Birds eat this dry berry, though we do not, and thus aid in dispersing the species. But though they digest the soft red outer pulp, formed by the swollen stalk, they cannot digest the hairy seed, so the plant attains its prime object of getting them duly scattered. The true roses, then, are another branch of the original potentilla stock, which have acquired a bushy mode of growth, with a fruit differing in construction from that of the brambles. Our English kinds are merely pink; the more developed exotics are often scarlet and crimson.
We have altogether some five true wild roses in Britain. The commonest is the dog-rose, which everybody knows well; and next comes the almost equally familiar sweet-briar, with its delicately scented glandular leaves. The burnet-rose is the parent of our cultivated Scotch roses, and the two other native kinds are comparatively rare. Double garden roses are produced from the single five-petalled wild varieties by making the stamens (which are the organs for manufacturing pollen) turn into bright-coloured petals. There is always more or less of a tendency for stamens[215] thus to alter their character; but in a wild state it never comes to any good, because such plants can never set seed, for want of pollen, and so die out in a single generation. Our gardeners, however, carefully select these distorted individuals, and so at length produce the large, handsome, barren flowers with which we are so familiar. The cabbage and moss roses are monstrous forms thus bred from the common wild French roses of the Mediterranean region; the China roses are cultivated abortions from an Asiatic species; and most of the other garden varieties are artificial crosses between these or various other kinds, obtained by fertilising the seed vessels of one bush with pollen taken from the blossoms of another of a different sort. To a botanical eye, double flowers, however large and fine, are never really beautiful, because they lack the order and symmetry which appear so conspicuously in the five petals, the clustered stamens, and the regular stigmas of the natural form.
From the great central division of the rose family, thus represented by the potentillas, the strawberry, the brambles, and the true roses, two main younger branches have diverged much more widely in different directions. As often happens, these junior offshoots have outstripped and surpassed the elder stock in[216] many points of structure and function. The first of the two branches in question is that of the plum-tribe; the second is that of the pears and apples. Each presents us with some new and important modifications of the family traits.
Of the plum tribe, our most familiar English examples, wild or cultivated, are the sloe or blackthorn, with its descendant the garden plum; as well as the cherry, the apricot, the peach, the nectarine, and the almond. All these plants differ more or less conspicuously from the members of the central group which we have so far been examining in their tree-like size and larger trunks. But they also differ in another important point: each flower contains only one seed instead of many, and this seed is inclosed in a hard bony covering, which causes the whole plum tribe (except only the almond, of which more anon) to be popularly included under the common title of ‘stone-fruits.’ In most cases, too, the single seed is further coated with a soft, sweet, succulent pulp, making the whole into an edible fruit. What, now, is the reason for this change? What advantage did the plant derive from this departure from the ordinary type of rose-flower and rose-fruit? To answer this question we must look at one particular instance in detail, and we cannot do better than take that well[217]-known fruit, the cherry, as our prime example of the whole class.
The cherry, like the strawberry, is an eatable fruit. But while in the strawberry we saw that the pulpy part consisted of the swollen stalk or receptacle, in which several small dry seeds were loosely embedded, with the cherry the pulpy part consists of the outer coat of the fruit or seed vessel itself, which has grown soft and juicy instead of remaining hard and dry. In this respect the cherry resembles a single grain from a raspberry; but from the raspberry, again, it differs in the fact that each flower produces only a single solitary one-seeded fruit, instead of producing a number of little fruits, all arranged together in a sort of thimble. In the raspberry flower, when blossoming, you will find in the centre several separate carpels or fruit-pieces; in the cherry you will find only one. The cherry, in fact, may (so far as its fruit is concerned) be likened to a raspberry in which all the carpels or fruit-pieces except one have become aborted. And the reason for the change is simply this: cherry bushes (for in a wild state they are hardly trees) are longer-lived plants than the bramble kind, and bear many more blossoms on each bush. Hence one seed to every blossom is quite as many as they require to keep up the numbers of the species.[218] Moreover their large and attractive fruits are much more likely to get eaten and so dispersed by birds than the smaller and less succulent berries of the brambles. Furthermore, the cherry has a harder stone around each seed, which is thus more effectually protected against being digested, and the seed itself consists of a comparatively big kernel, richly stored with food-stuffs for the young plant, which thus starts relatively well equipped in the battle of life. For all these reasons the cherries are better off than the brambles, and therefore they can afford to produce fewer seeds to each flower, as well as to make the coverings of these seeds larger and more attractive to birds. Originally, indeed, the cherry had two kernels in each stone, and to this day it retains two little embryo kernels in the blossom, one of which is usually abortive afterwards (though even now you may sometimes find two, as in philipœna almonds); but one seed being ordinarily quite sufficient for all practical purposes, the second one has long since disappeared in the vast majority of cases.
The plum scarcely differs from the cherry in anything important except the colour, size, and shape of the fruit. It is, as we have already noted, a cultivated variety of the blackthorn, in which the bush has become a tree, the thorns have been eradicated, and the[219] fruit has been immensely improved by careful selection. The change wrought in these two wild bushes by human tillage shows, indeed, how great is the extent to which any type of plant can be altered by circumstances in a very short time. The apricot is yet another variety of the same small group, long subjected to human cultivation in the East.
Peaches and nectarines differ from apricots mainly in their stones, which are wrinkled instead of being smooth; but otherwise they do not seriously diverge from the other members of the plum tribe. Indeed, though botanists rank the apricot as a plum, because of its smooth stone, and put the peach and nectarine in a genus by themselves, because of their wrinkled coating, common sense shows us at once that it would be much easier to turn an apricot into a peach than to turn a plum into an apricot. There is one species of nectarine, however, which has undergone a very curious change, and that is the almond. Different as they appear at first sight, the almond must really be regarded as a very slightly altered variety of nectarine. Its outer shell or husk represents the pulpy part of the nectarine fruit; and indeed, if you cut in two a young unripe almond and a young unripe nectarine, you will find that they resemble one another very closely. But as they ripen the outer coat of the nectarine[220] grows juicier, while that of the almond grows stringier and coarser, till at last the one becomes what we commonly call a fruit, while the other becomes what we commonly call a nut. Here, again, the reason for the change is not difficult to divine. Some seeds succeed best by making themselves attractive and trusting to birds for their dispersion; others succeed best by adopting the tactics of concealment, by dressing themselves in green when on the tree, and in brown when on the ground, and by seeking rather to evade than to invite the attention of the animal world. Those seed vessels which aim at the first plan we know as fruits; those which aim rather at the second we know as nuts. The almond is just a nectarine which has gone back to the nut-producing habit. The cases are nearly analogous to those of the strawberry and the potentilla, only the strawberry is a fruit developed from a dry seed, whereas the almond is a dry seed developed from a fruit. To some extent this may be regarded as a case of retrogressive evolution or degeneration.
The second great divergent branch of the rose family—that of the pears and apples—has proceeded towards much the same end as the plums, but in a strikingly different manner. The apple kind have grown into trees, and have produced fruits. Instead,[221] however, of the seed vessel itself becoming soft and succulent, the calyx or outer flower covering of the petals has covered up the carpels or young seed vessels even in the blossom, and has then swollen out into a sort of stalk-like fruit. The case, indeed, is again not unlike that of the strawberry, only that here the stalk has enlarged outward round the flower and inclosed the seeds, instead of simply swelling into a boss and embedding them. In the hip of the true roses we get some foreshadowing of this plan, except that in the roses the seeds still remained separate and free inside the swollen stalk, whereas in the pear and apple the entire fruit grows into a single solid mass. Here also, as before, we can trace a gradual development from the bushy to the tree-like form.
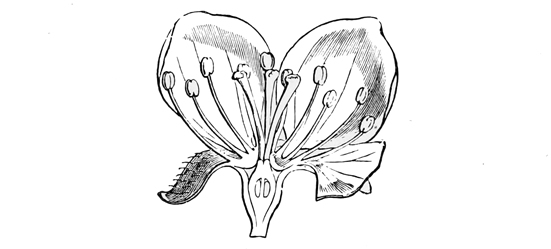
Fig. 47.—Vertical section of Apple-blossom.
The common hawthorn of our hedges shows us, perhaps, the simplest stage in the evolution of the apple tribe. It grows only into a tall bush, not[222] unlike that of the blackthorn, and similarly armed with stout spines, which are really short sharp branches, not mere prickly hairs, as in the case of the brambles. Occasionally, however, some of the hawthorns develop into real trees, with a single stumpy trunk, though they never grow to more than mere small spreading specimens of the arboreal type, quite unlike the very tall and stately pear-tree. The flowers of the hawthorn—may-blossom, as we generally call them—are still essentially of the rose type; but, instead of having a single embryo seed and simple fruit in the centre, they have a compound fruit, inclosing many seeds, and all embedded in the thick fleshy calyx or flower-cup. As the haw ripens the flower-cup outside grows redder and juicier, and the seed pieces at the same time become hard and bony. For it is a general principle of all edible fruits that, while they are young and the seeds are unripe, they remain green and sour, because then they could only be losers if eaten by birds; but as the seeds ripen and become fit to germinate, the pulp grows soft and sweet, and the skin assumes its bright hue, because then the birds will be of service to it by diffusing the mature seeds. How largely birds assist in thus dispersing plants has very lately been proved in Australia, where a new and troublesome weed has[223] rapidly overrun the whole country, because the fruit-eaters are very fond of it, and scatter its seeds broadcast over the length and breadth of the land.
The common medlar is nothing more than a hawthorn with a very big overgrown haw. In the wild state it bristles with hard thorns, which are wanting to the cultivated form, and its flower almost exactly resembles that of the may. The fruit, however, only becomes edible after it begins to decay, and the bony covering of the seeds is remarkably hard. It seems probable that the medlar, originally a native of southern Europe, is largely dispersed, not by birds, but by mice, rats, and other small quadrupeds. The colour is not particularly attractive, nor is the fruit particularly tempting while it remains upon the bush; but when it falls upon the ground and begins to rot, it may easily be eaten by rodents or pigs, and thus doubtless it procures the dispersion of its seeds under conditions highly favourable to their proper growth and success in life.
The little Siberian crabs, largely cultivated for their fruit in America, and sometimes found in English shrubberies as well, give us one of the earliest and simplest forms of the real apple group. In some respects, indeed, the apples are even simpler than the hawthorn, because their seeds or pips are not[224] inclosed in bony cases, but only in those rather tough leathery coverings which form what we call the core. The haw of the hawthorn may be regarded as a very small crab-apple, in which the walls of the seed cells have become very hard and stony; or the crab may be regarded as a rather large haw, in which the cell walls still remain only thinly cartilaginous. The flowers of all the group are practically identical, except in size, and the only real difference of structure between them is in the degree of hardness attained by the seed covers. The crabs, the apples, and the pears, however, all grow into tallish trees, and so have no need for thorns or prickles, because they are not exposed to the attacks of herbivorous animals. Ordinary orchard apples are, of course, merely cultivated varieties of the common wild crabs. In shape the apple-tree is always spreading, like an arboreal hawthorn, only on a larger scale. The pear-tree differs from it in two or three small points, of which the chief are its taller and more pyramidal form, and the curious tapering outline of the fruit. Nevertheless, pear-trees may be found of every size and type, especially in the wild state, from a mere straggling bush, no bigger than a hawthorn, to a handsome towering trunk, not unlike an elm or an alder.
In the matter of fruits, the apple group are more[225] advanced than the roses, but so far as regards the flower alone, viewed as an organ for attracting insects, many of the apple tribe are inferior to the true roses. Here again, however, we can trace a regular gradation from the small white blossoms of the may, through the larger blushing pink flowers of the apple, to the very expanded and brilliant crimson petals of that beautiful ornamental species of pear, the Pyrus japonica, so often trained on the sunny walls of cottages.
The quince is another form of apple very little removed from its congeners except in the fruit. More different in external appearance is the mountain-ash or rowan-tree, which few people would take at first sight for a rose at all. Nevertheless, its flowers exactly resemble apple-blossom, and its pretty red berries are only small crabs, dwarfed, no doubt, by its love for mountain heights and bleak windy situations, and clustered closely together into large drooping bundles. For the same reason, perhaps, its leaves have been split up into numerous small leaflets, which causes it to have been popularly regarded as a sort of ash. In the extreme north, the rowan shrinks to the condition of a stunted shrub; but in deep rich soils and warmer situations it rises into a pretty and graceful tree. The berries are eagerly eaten by birds, for whose attraction most[226] probably they have developed their beautiful scarlet colour.
So far, all the members of the rose family with which we have dealt have exhibited a progressive advance upon the common simple type, whose embodiment we found in the little wayside potentillas. Their flowers, their fruits, their stems, their branches, have all shown a regular and steady improvement, a constant increase in adaptation to the visits of insects or birds, and to the necessities for defence and protection. I should be giving a false conception of evolution in the roses, however, if I did not briefly illustrate the opposite fact of retrogressive development or degeneration which is found in some members of the class; and though these members are therefore almost necessarily less familiar to us, because their flowers and fruits are inconspicuous, while their stems are for the most part mere trailing creepers, I must find room to say a few words about two or three of the most noteworthy cases, in order to complete our hasty review of the commonest rosaceous tribes. For, as we all know, development is not always all upward. Among plants and animals there are usually some which fall behind in the race, and which manage nevertheless to eke out a livelihood for themselves in some less honourable and[227] distinguished position than their ancestors. About these black sheep of the rose family I must finally say a few words.
In order to get at them, we must go back once more to that simple central group of roses which includes the potentillas and the strawberry. These plants, as we saw, are mostly small trailers or creepers among grass or on banks; and they have little yellow or white blossoms, fertilised by the aid of insects. In most cases their flowers, though small, are distinct enough to attract attention in solitary arrangement. There are some species of this group, however, in which the flowers have become very much dwarfed, so that by themselves they would be quite too tiny to allure the eyes of bees or butterflies. This is the case among the meadow-sweets, to which branch also the spiræas of our gardens and conservatories belong. Our common English meadow-sweet has close trusses of numerous small whitish or cream-coloured flowers, thickly clustered together in dense bunches at the end of the stems; and in this way, as well as by their powerful perfume, the tiny blossoms, too minute to attract attention separately, are able to secure the desired attentions of any passing insect. In their case, as elsewhere, union is strength. The foreign spiræas cultivated in our hothouses have even smaller[228] separate flowers, but gathered into pretty, spiky antler-like branches, which contrast admirably with the dark green of the foliage, and so attain the requisite degree of conspicuousness. This habit of clustering the blossoms which are individually dwarfed and stunted may be looked upon as the first stage of degradation in the roses. The seeds of the meadow-sweet are very minute, dry, and inedible. They show no special adaptation to any particular mode of advanced dispersion, but trust merely to chance as they drop from the dry capsule upon the ground beneath.
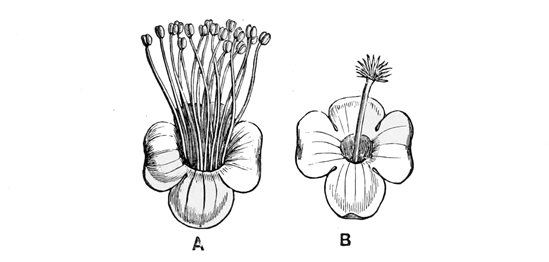
Fig. 48.—Single flower of Salad Burnet, male and female.
A far deeper stage of degradation is exhibited by the little salad-burnet of our meadows, which has lost the bright petals of its flowers altogether, and has taken to the wasteful and degenerate habit of fertilisation by means of the wind. We can understand the salad-burnet better if we look first at common agrimony, another little field weed about a foot high, with which most country people are familiar; for, though agrimony is not itself an example of degradation, its arrangement leads us on gradually to the lower types. It has a number of small yellow flowers like those of the cinquefoil; only, instead of standing singly on separate flower stalks, they are all arranged together on a common terminal spike, in the same way as in a hyacinth or a gladiolus. Now, agrimony is fertilised by[229] insects, and therefore, like most other small field roses, it has conspicuous yellow petals to attract its winged allies. But the salad-burnet, starting from a somewhat similar form, has undergone a good deal of degradation in adapting itself to wind-fertilisation. It has a long spike of flowers, like the agrimony; but these flowers are very small, and are closely crowded together into a sort of little mop-head at the end of the stem. They have lost their petals, because these were no longer needed to allure bees or butterflies, and they retain only the green calyx or flower-cup, so that the whole spike looks merely a bit of greenish vegetation, and would never be taken for a blossoming head by any save a botanical eye. The stamens hang out on long thread-like stems from the cup, so that the wind may catch the pollen and waft it to a neighbouring head; while the pistils which it is to fertilise have[230] their sensitive surface divided into numerous little plumes or brushes, so as readily to catch any stray pollen grain which may happen to pass their way. Moreover, in each head, all the upper flowers have pistils and embryo seed vessels only, without any stamens; while all the lower flowers have stamens and pollen bags only, without any pistils. This sort of division of labour, together with the same arrangement of seed-bearing blossoms above and pollen-bearing blossoms below, is very common among wind-fertilised plants, and for a very good reason. If the stamens and pistils were inclosed in a single flower they would fertilise themselves, and so lose all the benefit which plants derive from a cross, with its consequent infusion of fresh blood. If, again, the stamens were above and the pistils below, the pollen from the stamens would fall upon and impregnate the pistils, thus fertilising each blossom from others on the same plant—a plan which is hardly better than that of self-fertilisation. But when the stamens are below and the pistils above, then each flower must necessarily be fertilised by pollen from another plant, which ensures in the highest degree the benefits to be derived from a cross.
Thus we see that the salad-burnet has adapted itself perfectly to its new mode of life. Yet that[231] adaptation is itself of the nature of a degradation, because it is a lapse from a higher to a lower grade of organisation—it is like a civilised man taking to a Robinson Crusoe existence, and dressing in fresh skins. Indeed, so largely has the salad-burnet lost the distinctive features of its relatives, the true roses, that no one but a skilled botanist would ever have guessed it to be a rose at all. In outer appearance it is much more like the little flat grassy plantains which grow as weeds by every roadside; and it is only a minute consideration of its structure and analogies which can lead us to recognise it as really and essentially a very degenerate and inconspicuous rose. Yet its ancestors must once have been true roses, for all that, with coloured petals and all the rosaceous characteristics, since it still retains many traces of its old habits even in its modern degraded form.
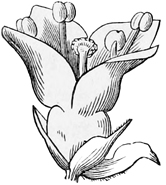
Fig. 49.—Flower of Stanch-Wound or Great Burnet.
We have in England another common weed, very like the salad-burnet, and popularly known as stanch-wound, or great-burnet, whose history is quite as interesting as that of its neighbour. The stanch-wound is really a salad-burnet which has again lost its habit of depending upon the wind for fertilisation, and has reverted to the earlier insect-attracting tactics of the race. As it had already lost its petals, how[232]ever, it could not easily replace them, so it has coloured its calyx or flower-cup instead, which answers exactly the same purpose. In other words, having no petals, it has been obliged to pour the purple pigment with which it allures its butterfly friends into the part answering to the green covering of the salad-burnet. It has a head of small coloured blossoms, extremely like those of the sister species in many respects, only purple instead of green. Moreover, to suit its new habits, it has its cup much more tubular than that of the salad-burnet; its stamens do not hang out to the wind, but are inclosed within the tube; and the pistil has its sensitive surface shortened into a little sticky knob instead of being split up into a number of long fringes or plumes. All these peculiarities of course depend upon its return from the new and bad habit of wind-fertilisation to the older and more economical plan of getting the pollen carried from[233] head to head by bees or butterflies. The two flowers grow also exactly where we should expect them to do. The salad-burnet loves dry and wind-swept pastures or rocky hill-sides, where it has free elbow-room to shed its pollen to the breeze; the stanch-wound takes rather to moist and rich meadows, where many insects are always to be found flitting about from blossom to blossom of the honey-bearing daisies or the sweet-scented clover.
Perhaps it may be asked, How do I know that the salad-burnet is not descended from the stanch-wound, rather than the stanch-wound from the salad-burnet? At first sight this might seem the simpler explanation of the facts, but I merely mention it to show briefly what are the sort of grounds on which such questions must be decided. The stanch-wound is certainly a later development than the salad-burnet; and for this reason—it has only four stamens, while the parent plant has several, like all the other roses. Now, it would be almost impossible for the flower first to lose the numerous stamens of the ordinary rose type, and then to regain them anew as occasion demanded. It is easy enough to lose any part or organ, but it is a very different thing to develop it over again. Thus the great-burnet, having once lost its petals, has never recovered them, but has been obliged to colour[234] its calyx instead. It is much more natural, therefore, to suppose that the stanch-wound, with its few stamens and its clumsy device of a coloured calyx instead of petals, is descended from the salad-burnet, than that the pedigree should run the other way; and there are many minor considerations which tend in the same direction. Most correctly of all, we ought perhaps to say that the one form is probably a descendant of ancestors more or less like the other, but that it has lost its ancestors’ acquired habits of wind-fertilisation, and reverted to the older methods of the whole tribe. Still, it has not been able to replace the lost petals.
I ought likewise to add that there are yet other roses even more degenerate than the burnets, such as the little creeping parsley-piert, a mere low moss-like plant, clinging in the crannies of limestone rocks or growing on the top of earthy walls, with tiny green petalless flowers, so small that they can hardly be distinguished with the naked eye. These, however, I cannot now find space to describe at length; and, indeed, they are of little interest to anybody save the professional botanist. But I must just take room to mention that if I had employed exotic examples as well as the familiar English ones, I might have traced the lines of descent in some cases far more fully. It is perhaps better, however, to confine our attention[235] to fairly well-known plants, whose peculiarities we can all carry easily in our mind’s eye, rather than to overload the question with technical details about unknown or unfamiliar species, whose names convey no notion at all to an English reader. When we consider, too, that the roses form only one family out of the ninety families of flowering plants to be found in England alone, it will be clear that such a genealogy as that which I have here endeavoured roughly to sketch out is but one among many interesting plant pedigrees which might be easily constructed on evolutionary principles. Indeed, the roses are a comparatively small group by the side of many others, such as the pea-flowers, the carrot tribe, and the dead-nettles. Thus, we have in England only forty-five species of roses, as against over two hundred species of the daisy family. Nevertheless, I have chosen the rose tribe as the best example of a genealogical study of plants, because most probably a larger number of roses are known to unbotanical readers than is the case with any other similar division of the vegetable world.

Fig. 50.—Cuckoo-Pint (Arum Maculatum).
Close by the hedge-side there runs a little streamlet known to the village children for two miles around by the strangely pleonastic title of the Bourne Brook. Pleonastic, I say, because bourne is, of course, good old English for what in modern English we call a brook, so that the two halves of the common name are, in fact, synonymous, the later word being added to the earlier by the same sort of unconscious reduplication as that which gives us the double forms of Windermere Lake or Mount Ben Jerlaw. I can’t tell you, though, what a world of life and interest is to be found among the low cliff banks and tiny shingle patches that bound the Bourne Brook. In the stream itself there are darting crayfish, which we can catch with our fingers by lifting up the green slimy stones; there are caddis-worms, and big pond snails, and pouting miller’s thumbs, and iridescent stickleback; it is even rumoured, though I doubt whether on sufficient[237] authority, that there are actually and positively in some of its pools and stickles genuine unadulterated real live trout. I know as a fact, however, that there are freshwater mussels, for these I have fished up with my little dredging-net, and safely domesticated in the bell-glass aquarium. In the fields around there are ferns, and marsh-marigolds, and rushes, and roast-beef plants. And beside the water’s edge there are[238] abundant leaves and blossoms of that strange flower the cuckoo-pint, whose counterfeit presentment you see in the figure on the previous page. Now, cuckoo-pint, or lords-and-ladies, or wild arum, whichever you choose to call it, is a very singular plant indeed; and it seems to me we cannot do better than sit down and dissect one for the sake of understanding its queer internal arrangements. If it were a newly discovered Central African lily, we should all be reading about its extraordinary adaptations in all the newspapers: much more then, since it is a common English plant we have all known familiarly from childhood upward, ought we to wish for some explanation of its singular shape and its wonderful devices for entrapping and intoxicating helpless little flying insects.
First of all, we must begin by recognising that the apparent flower of the cuckoo-pint is not one single blossom, but a whole group of separate blossoms, closely crowded together in two or three little distinct bundles on a long spike or succulent stem. And in order to let us all clearly understand the meaning and nature of the entire compound structure, I think we had better divide our subject (as if it were a sermon) into three heads. First, we must consider what are the actual parts to be distinguished from one another in the flower of the cuckoo-pint at the present day.[239] Secondly, we must ask what was the course of evolution by which they each assumed those peculiar forms. And thirdly, we must inquire what good purpose in the economy of the plant is subserved by each part in the existing cuckoo-pints as we now find them. We shall thus have learnt, at last, what a cuckoo-pint is, how it came to be so, and why its various portions have been brought to assume their present forms.
Fig. 51.
Spadix of Arum.
Beginning, then, with the purely structural or positive arrangement of the cuckoo-pint as we find it in nature at the present day, we see at once that its blossom consists mainly of a large greenish-purple sheath or hood, at the top of a long stalk, inclosing a tall fleshy spike or club, shaped something like a mace, and protruding from the hood in front, so as to show its coloured and expanded summit above the point of junction of the two lips. That is all that one can see of the blossom from the outside; but in reality these two conspicuous organs form no part of the actual and genuine flowers themselves at all. They are merely incidental accessories, put there for an excellent pur[240]pose indeed, as everything always is in the balanced economy of nature; but not essential or necessary to the existence of the flowers as flowers, though most noticeable from their size and hue to the superficial eyes of the unscientific human kind. In order to see the true flowers themselves, we must cut open the side of the hood or sheath, as has been done in the accompanying diagram, and then one can observe a number of small knobby bodies clustering in three groups along the lower part of the club-shaped spike or central axis. Those little knobby bodies, of which there are a great many in each arum, form the real blossoms of the cuckoo-pint; and they are inclosed in the sheathing hood for a very good reason, as we shall hereafter see, in order to ensure the carrying out of their proper function, the final production of seeds and berries.
If one looks closely at the diagram, however, one can notice that these little knobby flowers are not all quite similar to one another. They consist of three distinct kinds, all three of which are always found in true arums of this type. At the bottom there are a whole group of small cushion-like green lumps, each with a little point in its centre, and all closely packed together in several irregular rows, like Indian corn on the cob. These green lumps are the pistil-bearing[241] flowers; each of them represents a single very degraded blossom, and each will grow out at a later stage into one of the bright scarlet berries which form such beautiful objects in the hedgerow and waysides during the autumn months. We could not possibly have a simpler type of flower than these lowest pistil-bearing blossoms; they are in fact the central floral notion reduced to its ideally simplest terms. They consist each of a single rudimentary berry, containing a single seed, and crowned by a little point or stigma, which is the sensitive surface to be fertilised by the pollen from the other flowers.
In the middle, here, come the flowers of this second or pollen-bearing sort, each of which again consists of naked stamens; that is to say, each flower is here reduced to one solitary part, analogous to the little pollen-sacs that you see hanging out in the centre of a tiger lily or most other conspicuous garden blossoms. Every such stamen is made up of two tiny bags, which open when ripe and discharge their golden pollen. Though the pollen looks to the naked eye like mere yellow dust, yet, when put under a microscope, it is seen to consist of small egg-like bodies, having a characteristic shape and appearance in each different flower, exactly as the seeds and fruits have to the naked eye. With these two essential[242] elements of floral architecture we are now pretty familiar from our previous examination of other plants.
On top of all, however, come a group of very peculiar blossoms, found only in the arum and nowhere else, and consisting of several little green knobs, like those of the pistil-bearing flowers, but each crowned by a long hair or filament, bent downwards towards the base of the hood or sheath, and very much larger than the sensitive surface of the lower blossoms. The origin and meaning of these peculiar organs we will come to consider later on: for the present it will be sufficient to observe their shape and position, and to notice that their hairs point downward and inward like the spikes of a lobster pot, at a point exactly corresponding to the narrowest neck or throat of the inclosing sheath.
And now, how did the cuckoo-pint come to possess this very singular arrangement of tiny separate flowers in a close spike, female below, male in the centre, and neuter or rudimentary on the top of all? To answer this question properly, we must go back to the earlier ancestors of the arum tribe—and I may as well start fair by saying at once that the arums are by descent degenerate lilies, like wheat, and that each of these very degraded little flowers really represents a primitive full-blown and bright-coloured lily blossom. You[243] will remember that a true lily is made up of six brilliant petals or flower-leaves, inclosing six long pendulous stamens, and with a seed-vessel or ovary of three cells in the very centre. Such a blossom as that we call a perfect flower, because it possesses within itself all the component elements of any blossom—calyx, petals, stamens, and pistil. Moreover, it is, so to speak, a self-contained and self-sufficing flower; it has bright petals to entice an insect fertiliser, pollen to impregnate its ovary, and embryo seeds to form the future ripe fruit. But as we have so often noticed, it is highly undesirable for a flower to be fertilised with pollen from its own stamens: those plants which are impregnated from the stamens of their neighbours always produce more seed and stronger seedlings than those which are impregnated with home-made pollen from their own sacs. Hence, cross-fertilisation, we know, is the great end aimed at by all flowers; and those plants which happen to vary in any direction favourable to cross fertilisation invariably succeed best in the struggle for life, while those which happen to vary in any direction hostile to it, or which acquire the bad habit of self-fertilisation, tend slowly to go to the wall and to die out from inherited and ever-increasing feebleness of constitution.
There can be very little doubt that the ancestors[244] of the arums had originally six coloured petals like the lilies, for a reason which I will shortly mention; and inside these petals were six stamens and a three-celled ovary or unripe capsule. It is a very long step, certainly, from such perfect flowers as those to such very rudimentary and reduced types as the little florets which we get in the cuckoo-pint, each consisting of a few stamens or a single one-seeded fruitlet, without any trace of petals whatsoever. Yet we have very good evidence of the slow course of degradation by which the arums have reached their present condition; and, as happened in the case of wheat, several surviving intermediate forms enable us to bridge over the intervening gulf. In other words, there are plants which resemble the lilies in some things, while they resemble the arums in others; and by means of these plants we can trace a regular gradation from the perfect and bright-coloured flowers of the true lily to the imperfect and inconspicuous little unisexual blossoms of our English cuckoo-pint. It is interesting, too, to observe how the very same original stock which in one direction gave birth to the degenerate wind-fertilised wheat and grasses, has in another direction given birth to the equally degenerate but insect-fertilised arums and their congeners. The one case shows the course of degradation as it takes place in poor dry soils; the other[245] case shows it as it takes place in the moist and rich mould of watery ditches.
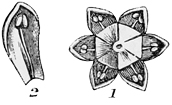
Fig. 52.—Single flower of Sweet-sedge.
Look first at the curious flower which is represented for us here in the little sketch at the side. In the slow rivers of Suffolk, and along the shallow edges of the Norfolk broads, there grows a pretty spiky water-plant, known by the scientific name of Acorus, or by the simpler English titles of sweet-flag and sweet-sedge. This acorus is a highly aromatic reed-like plant, with long lance-shaped leaves, and a dense spike of small yellowish-green blossoms, standing out in a cylindrical form from the thick rod which does duty for its stem. At first sight you would not say that these flowers differed very much from those of the arum: they look pretty much the same sort of small unnoticeable green knobs to a casual observer. But when one comes to pick out one of them from the close mass, and to examine it with a common pocket lens, one can see at once that, though very much reduced in size and colour, it is still at bottom essentially a lily flower. In the diagram we have one of these small blossoms considerably enlarged, and it is easy to see that it possesses all the various parts which characterise the true lilies. There are six petals, clearly enough,[246] though they are minute and green instead of being brilliantly coloured; and they are closely folded over the central organs, instead of being bent back and displayed ostentatiously to the eyes of passing insects. There are six stamens too, one under each petal, almost concealed by the scale-like covering; and in the centre there is an ovary which when cut across proves to have sometimes two and sometimes three seed-bearing cells, for the number here has become a little indefinite: nature, as so often happens, has begun to lose count. There can be no sort of doubt, then, that acorus represents a very reduced and degraded lily, still retaining all its primitive structural arrangements, but with its flowers greatly diminished in size, and with its original bright colour almost entirely lost by disuse and degradation.
The reason why this little acorus or sweet-sedge has thus gone backward in the course of development is not a very difficult one to understand. Brilliant flowers like the lilies depend for fertilisation upon large colour-loving insects, such as bees and butterflies, which are attracted by their flaunting hues and their abundant store of rich honey, and so unconsciously carry the impregnating pollen from head to head. But many other plants find it suits their purpose better to depend either upon the wind or upon[247] small insect friends of less pronounced æsthetic tastes; and this is especially the case, among other classes, with almost all waterside plants. Hence such plants have usually acquired small and inconspicuous separate flowers; and then, to make up for their loss in attractiveness, like cheap sweetmeats, they have very largely increased their numbers. Or, to put the matter more simply and physically, in waterside situations those plants succeed best which have a relatively large number of individually small and unnoticeable flowers, massed together into large and closely serried bundles. Hence, in such situations, there is a tendency for petals to be suppressed, and for blossoms to grow minute; because the large and bright flowers seldom succeed in attracting big land-insects like bees or butterflies, while the small and thick-set ones usually do succeed in attracting a great many little flitting waterside midges. Examples may be found in the rushes, bur-reeds, catstails, and many other freshwater plants.
For such a rôle our friend the sweet-sedge is peculiarly well adapted. Its small yellowish blossoms, though separately unnoteworthy, are rendered conspicuous in the mass by their dense grouping: and its extremely aromatic perfume makes it a great favourite with the tiny flies and water-haunting insects, who[248] are much more guided in their search for food by scent than by sight. These little flies carry its pollen from one head to another, and so unconsciously fertilise the future seeds, and give the plant a firm foothold in all situations which are naturally suitable for its peculiar mode of growth.
Fig. 53.—Single flower of
Marsh Calla.
The common marsh calla of northern Europe (fig. 53) bridges over the gap between this English plant and the stages below it on the path of degradation. Calla has by disuse quite lost its petals, but it nevertheless retains six stamens to each flower, grouped round a single ovary. Here the close relationship to the true lilies still remains quite apparent.
Next in descending order, on the way to the cuckoo-pint, we may take that common white lily which grows so often in cottage windows, and which boasts more names, Latin and English, than almost any other plant whose personal acquaintance I have ever had the pleasure of making. The members of a Sheffield long firm themselves have seldom so many aliases as this honest and unoffending flower. Botanists call it Richardia Africana; gardeners dub it Calla Æthiopica; and the general public knows it[249] indiscriminately as Æthiopian lily, white calla, snowy arum, St. Helena arrowroot, and lily of the Nile. However, in spite of its numerous disguises, I dare say it will be easy to recognise the plant I mean, when I say that it is very much like a cuckoo-pint, only with a pure snow-white hood, and a bright golden yellow spike projecting from the top. As in the cuckoo-pint this golden spike is the part which contains the true flowers; and the snow-white hood is only a sort of shroud or cloak which covers them in from the vulgar gaze. The Æthiopian lily, then (since we must choose one among its many names), presents us with a further step on the downward path of degradation from the true lilies towards the thoroughgoing cuckoo-pint: for, as preachers justly remark, there is no drawing a line after you have once begun upon the wrong track, and a lily which lets in the thin end of the wedge by becoming a sweet-sedge is almost certain to end at last, in the form of its remote descendants, as a mere degenerate and neglected arum.
When we cut open the hood of the Æthiopian lily, we find inside it a spike somewhat resembling that of the cuckoo-pint, but differing in one or two important particulars. Near the bottom, at a point corresponding to that where the female flowers grow[250] in the wild English arum, the white Æthiopian lily has a number of small greenish knobs, apparently embedded in a golden yellow matrix; at the top, the whole of the spike consists of a similar golden-yellow substance, which, at a certain period in the flowering process, effloresces, so to speak, with a copious greasy white dust, something like starch or wheaten flour. But if we split down the spike itself through the centre, we can soon find out what is the meaning of this curious arrangement. The golden substance which makes up the mass of the spike consists really of innumerable yellow stamens, packed so tightly together over the whole stem, and so closely sessile (as we call it technically) upon the central axis, that they look like a single piece of homogeneous waxy material. You can separate them from one another, however, with your fingers, and then you see that each one is roughly pentagonal or hexagonal in outline, owing to the pressure of its surrounding neighbours, and that it consists essentially of a small pollen bag, containing a quantity of yellowish liquid. When the stamens are quite ripe, this liquid assumes the form of small white pollen grains, which are pushed out as the bags open, and become the efflorescence or powder that covers the spike in its ripe state. At the bottom of the spike, where we get the[251] pistil-bearing flowers in the cuckoo-pint, the Æthiopian lily has several small blossoms intermediate between the perfect flowers of the acorus, or the half-perfect flowers of the marsh calla, and the very imperfect flowers of the arum; for each of them has here a central green knob or capsule, surrounded irregularly by four or five stamens, but without any petals, or even any scales to represent them. These form the green bodies which I have already described as apparently embedded in a hard yellow matrix; and that yellow matrix is composed of the stamens. The lower part of the Æthiopian lily, in fact, consists of irregular flowers which, like those of the marsh calla, have quite lost their petals, but which still retain an indefinite number of stamens grouped around a single pistil; while in the upper part, as in the central group of the arum, the pistils have disappeared also, and only the stamens remain. Such a plant as this lily, then, is clearly on the way to becoming what the arum has actually become: its flowers already show a tendency toward the unisexual condition. In the upper portion they have all become actually unisexual, for there we get nothing but stamens; in the lower part they remain irregularly bisexual, for there, though the stamens are often reduced in number—nature losing count again—some of them still remain embedded[252] on the spike between the scattered pistils. This result is just what you might naturally expect from squeezing a lot of marsh calla blossoms closely together on a spike. Even in the upper half of the spike, the blossoms often keep up some marks of their original bisexual character, for you will occasionally find a few stray green knobs sparsely sprinkled here and there among the golden stamens of the top portion. Nevertheless, we may fairly say that even here a tendency towards specialisation has been distinctly set up: the uppermost flowers tend to become almost entirely pollen bearing sacs, and the lowermost flowers tend to become preponderatingly, though not entirely, seed-bearing ovaries.
Now if we turn from these transitional steps to the completely developed arum, what do we find? Here, the top of the spike has become absolutely bald and bare of flowers, instead of being covered, as in the Æthiopian lily, with thickly grouped florets up to its very summit; and at the same time, the actual flowers in the lower portion, instead of running together into an uninterrupted cone, are separated into three distinct groups or bodies. At the bottom of all, as in the Æthiopian lily, we now get the female flowers alone; only, instead of being intermixed with stamens, they consist simply of naked ovaries; the[253] differentiation or specialisation of the flowers is here complete. Above them, as before, we get the male flowers, reduced to a single stamen, or rather to a group of from four to six stamens each, all run together: for though it is usual to consider each stamen as a separate flower (which it certainly is in some still more degraded arums, like the little ‘Capuchin’ of southern Europe), I think the analogy of marsh calla and the Æthiopian lily justifies us in regarding them as groups of six, more or less defective, and jammed closely together, with the ovaries crushed out between them. And at the top of all we get a perfectly new factor in the compound community—a number of green sacs capped by downward-pointing hairs, which are, in fact, abortive pistils, like those organs that form the lower group, only with their ovaries barren, and their styles or sensitive surfaces lengthened out into spiky hairs.[11] What may be the use or function of these curious objects we will proceed to inquire a little later: for the present we must turn our attention to the origin of another part of the cuckoo-pint’s apparent blossom, the large and conspicuous greenish-purple hood, which alone[254] composes the flower in the popular sense of the word.
There is nothing at all like that, a casual observer would probably be tempted at first to say, in any ordinary true lily that any one ever yet came across. A bunch of lilies growing on a stalk, with a sort of huge winding-sheet wrapped round them, is a thing that surely nobody has ever seen. So it would seem at a first glance; and yet there is one lily-like plant that we all know well, in which the flowers are at one time wrapped up in exactly such an enveloping sheet. Have you ever watched a narcissus or a daffodil unfolding its pretty yellow buds? If you have, you will remember that at first they are all tightly covered over by a thin papery membrane, shaped exactly like the hood of this cuckoo-pint; and that after the scented blossoms have all come out, this membrane, or spathe, as we call it in the horrid technical language of botany, turns back upon the stem, like a sort of cup below the flowers. To be sure, the daffodil and the narcissus are not, in the strictest sense of the word, true lilies at all, but amaryllids, because they have got an inferior instead of a superior ovary; but, even in the technically restricted lily family itself, there are lilies with just such a spathe or enveloping membrane, as in the familiar head of onions and garlic, as well as in some[255] more respectable and dignified flowers. Now, one has only got to suppose the number of buds in each head largely increased, the whole head lengthened out into a spike, and the spathe or sheath grown larger into a completely inclosing hood, and there we have at once an arum or an Æthiopian lily. Only, as often happens under such circumstances, the individual flowers have now grown too small to attract the fertilising insects separately on their own account; so the spathe or hood has to do duty for them all at once collectively. It incloses and conceals the various minute flowers, but it becomes itself coloured and attractive, so as to allure the eyes of the little insects on behalf of the entire community. In other words, when the central flowers had become so much diminished in size by disuse, by loss of their petals, and by specialisation of sexes, they ran no chance of getting fertilised at all unless they possessed some exceptional means of attracting insects. Hence those alone have survived which happened to develop some such attractive organ as the hood of the Æthiopian lily or the purple central spike of the English arum.
And now we come at last to the final purpose of all these curious structural arrangements. The object of them all is of course to ensure the cross-fertilisation of the different heads of flowers; but the special way in which they effect this universal end is singularly[256] ingenious, interesting, and almost intentional in its design. The Æthiopian lily, one can readily understand, attracts many insects by its large brilliant white hood, as well as by the rich golden-yellow colour of the stamens which cover the summit of its spike. But in the arum the top of the spike is bare, and has become expanded into a club-shaped organ, which is deeply tinged with purple, and stands out vividly against the bright green of the spathe at its back, so as to form an excellent advertisement for the giddy eyes of little passing winged insects. It is upon these insects that the arum depends entirely for fertilisation, and the way in which it manages to obtain their services is as curious as anything in the whole range of vegetable existence.
If, when the arum-flowers are just beginning to blossom, I were to cut down one of the hoods halfway through the centre, sideways, I should find a great many tiny winged flies all crawling about at the very bottom of the deep tube. They have come from some other neighbouring arum-flower, where they have been well dusted with the golden pollen; and they crawl down the neck of the hood, past the lobster pot hairs which close its narrowest portion, into the broader open space beneath. Here they find the pistil-bearing blossoms just ripe for impreg[257]nation; and, crawling over them in an aimless sort of fashion, they rub off upon their sensitive surfaces some of the pollen which they brought with them from the last plant they visited. This pollen thus cross-fertilises the fruit, and produces in it seeds which are the product of two distinct parents, and therefore capable of springing up into vigorous seedlings of the strongest sort.
But though the small flies have thus benefited the plant by fertilising its ovaries with pollen brought from another head, they have as yet got no return for their trouble in the shape of meat or drink: and, unless they did so, they certainly would not take the trouble to visit any other flower of the same sort. The stamens are not yet ripe, and do not ripen until after the pistils have set their fruit. If they did otherwise, then the pollen would fall from them down upon the sensitive surfaces of their sister blossoms below, and the plant would accordingly be self-fertilised—a thing to be always avoided as far as possible. Accordingly, it is a fixed rule in the cuckoo-pints that the pistils, which are below, come to maturity first, while the stamens, which are above, shed their stock of pollen a day or two later. This being so, the flies find nothing in the new flower to detain them any longer; and, if they could, they[258] would crawl up the spike and get out again by the same way as they got in, never troubling themselves any more about such useless flowers. Here, however, the curious lobster-pot hairs for the first time come into play. They act, in short, exactly like a common eel-trap. The flies walked in easily enough, the way the hairs naturally pointed; but when they try to walk out again, they find their way completely blocked by the chevaux-de-frise of stiff bristles. There is nothing that beats a crawling insect like a thicket of hairs; he finds it as impossible to creep up against their grain as we ourselves find it to force our way through a tropical jungle of cactus and prickly spurges. So there they wait perforce for a time in durance vile, wandering up and down helplessly among the lower flowers, and effectually brushing off against them every single grain of pollen which they brought on their legs or breasts from the last flower they visited.
At last, in a day or so, the young berries begin to swell slowly, and all the pistil-bearing flowers show by this quickening action that they have been duly and properly fertilised. Then comes the turn of the stamens. One after another they open their little double pollen-sacs, and shed their golden powder down upon the wings and bodies of the small flies[259] imprisoned beneath. Even if a little of it happens to catch upon the pistils here and there, that does not matter now, for all the ovaries are already duly impregnated, and the sensitive surfaces have shrivelled utterly away; so most of the pollen falls on to the floor of the hood, where the small flies are waiting impatiently and hungrily for the Danae flood. It covers them all over from head to foot with the golden grains, and clogs their legs and wings and bodies in every portion. A fine time the flies have of it then. They get actually drunk with pollen after their fast; and, if you cut open one of the hoods in this stage of development, you will find the little creatures positively reeling about in their intoxication, and so full-fed with rich grains that they can hardly use their legs or wings to crawl or fly. A little fresh air seems to revive them slightly, as is often the case with other gentlemen under similar circumstances; and then they can feebly fly away after a few minutes.
But in the natural state of things, when no wandering botanist comes with his penknife to make what he calls in his lively language a ‘longitudinal section of Arum maculatum,’ the flies remain at the bottom of their deep well till they have eaten almost all the pollen, and got most helplessly and stupidly drunk in the process. A great waste of pollen this, for the[260] plant, of course; but still it costs no more than honey would do, and quite enough remains on the legs and wings of the flies to impregnate their fellow-blossoms on another plant. At last all the pollen is shed and eaten, and then the flies again become anxious to shift their quarters to some more favourable spot, where there is more food to be found, and another drunken orgy to be expected. This time, however, the hairs no longer impede their progress; they have all shrivelled up meanwhile, and the eel-trap is therefore now dissolved; so the flies hurry away once more, covered with the stock of pollen-dust which has been showered down upon them by their late host.
One might suppose, at first, that after one such experience the flies would studiously avoid cuckoo-pints in future. Nothing of the sort. Experience seems to be thrown away upon insects; and besides, the little creatures seem actually to enjoy their intoxicated revels. Pollen apparently acts upon them as an incentive, exactly as opium acts upon a Chinaman. The first thing they do the moment they are released is to forthwith fly off to the nearest other cuckoo-pint. They see a purple, club-shaped spike, somewhere close by, overtopping the folded lips of the green hood, and they make straight for that well-known signpost, as the lordly human race makes for the flaring lights of[261] a gilded public-house. Once more they crawl down the funnel-shaped tube; once more they pass the eel-pot hairs; and once more they rub off the pollen that clings to their legs and sides upon the sensitive surfaces of the lower flowers. For a while they have again to fast in their narrow prison; and then the stamens of the second arum open their pollen-sacs, and dust the greedy insects a second or third time with golden grains. So, throughout the whole flowering season of the arums, these little flies go about from head to head in constant relays, unconsciously benefiting the plant, while they are effecting their own hungry purpose in eating up the spare pollen. From the point of view of the insects, the only use of arums is to produce food and shelter for wandering flies; but from the point of view of the plant, the only use of insects is to act as common carriers for the conveyance of pollen from one head to another. Man, however, is far wiser and more expansive in his ideas about the economy of nature than either: according to him, the real, final end of all this beautiful and marvellous mechanism is to produce Portland arrowroot for starching his own civilised shirt-fronts, wristbands, and collars.
After the dissolute small flies have performed their function in the economy of the cuckoo-pint by thus[262] fertilising the small green ovaries, the plant begins to enter upon a fresh phase of existence. It has now no further use for its hood and its purple-topped spike, which have answered their purpose in attracting the insects; and therefore it gets rid of them in the same summary way in which mankind generally get rid of a faithful old horse, or a superannuated servant. The hood withers slowly away; the top of the spike, as far down as the base of the cluster of stamens, gradually decays; and at last you find nothing left but a bunch of rather shapeless green berries, elevated on a stiff, fleshy stalk, and with a scar at their bottom in the place where the hood used once to join on. As summer wears away the berries grow bigger and bigger, while at the same time they become redder and redder. At last, with the first approach of autumn, they appear as the bright cluster of coral-coloured berries, represented at the side, with which we are all so familiar in our September hedgerows.

Fig. 54.
What is the use of this new manœuvre? Well, it is not simply that common to most succulent fruits. Each of these bright red berries incloses a single hard nut-like seed. Its object is to attract the fruit-eating birds, the field-mice, and the other small animals, to eat it up whole. For this end, just as so many flowers[263] have bright-coloured petals to attract the eyes of insects, we know that fruits have bright-coloured pulpy coverings to attract the eyes of birds or mammals. And as the flowers put honey in their nectaries as an allurement for the bees, so the fruits put sugary juices in their pulp as an allurement for the robins and bullfinches. So far, the trick is just the ordinary plan of all fruit-bearers. The arum, however, has a still more cruel and insidious mode of procedure. Its berries are poisonous; and very often, I believe, they destroy the little birds that they have enticed by their delusive prettiness. Then the body of the murdered robin decays away, and forms a mouldering manure-heap, from which the young cuckoo-pint derives a store of fresh nutriment. I will not positively assert that it is for this reason the cuckoo-pint has acquired its poisonous juices; but I cannot help seeing that if any berry happened to show any tendency in such a direction, and so occasionally poisoned the creatures which eat it, it would thereby obtain an advantage in the struggle for existence, and would tend to increase the poisonous habit so far as it continued to obtain any further advantage by so doing. To some people this may seem grotesque; but the grotesqueness is in the facts of nature, not in the appreciation of their inevitable results. Poisonous berries are unquestion[264]ably useful to the plants which bear them; and, if we find their usefulness ridiculous, that is a peculiarity of our own sense of humour which in no way affects the abstract truth of the observation. It is impossible, in fact, that a plant should not benefit by having its berries poisonous, and so some plants must necessarily, in the infinite variability of nature, acquire the property of killing their friendly allies. It has been asked why the birds have not on their side learnt that the arum is poisonous. The very question shows at once an ingrained inability to understand the working of natural selection. Every bird that eats arum berries gets poisoned: but the other birds don’t hold a coroner’s inquest upon its body or inquire into the cause of death. Naturally the same bird never eats the berries twice, and so experience has nothing more to do with the matter than in the famous illogicality about the skinning of eels.
There are many other curious points of interest about the arum: there are the glossy arrow-headed leaves; there is the sharp, deterrent, pungent juice; the tall, succulent, biting stem; the thick, starchy, poisonous root-stock, where the plant lays by the store of nutriment it collects each summer for next spring’s flowering season. All these demand and repay the minutest and most careful study. But life is too[265] short for us to know even a cuckoo-pint to the very bottom; and so, perhaps, instead of turning aside to other subjects of interest in its structure and functions, it will be best to recapitulate afresh from an historical point of view the main steps in the evolution of the arum tribe at which we have already glanced.
Originally, the ancestors of the arum were a sort of lilies, with bright petals, and with six stamens and a three-celled ovary to each flower. They had also a papery spathe or hood, like the narcissus and the onion, at the base of their blossoms; and this spathe has been gradually modified into the green cap of the modern cuckoo-pint. Slowly the flowers became reduced in size, like those of acorus; and then they grew degraded in structure, till at last they entirely lost all their petals—a stage at which the lower flowers of the Æthiopian lily still remain. Next, the blossoms began to differentiate into three distinct groups, which owed their specialised form to the new mode of insect fertilisation. The lowest flowers lost all their stamens, and were reduced to a single ovary each. The middle flowers lost all their ovaries, and were reduced to a few stamens each. The topmost flowers underwent a still more curious change, and after losing their stamens made their ovaries abortive, in order to act as eel-traps for the fertilising flies. The series of[266] alterations by which these structural modifications were brought about must have been very slow; and they must have been produced by the constant fertilisation of such arums as best retained the visiting flies, and the dying out of such as did not well retain them. Last of all, the berries grew large and red under the influence of animal selection, those berries which attracted birds succeeding in producing new plants, while those which did not so attract them died out unsuccessfully. And at the same time the ovary came to contain only one seed, instead of three cells with many seeds, because one seed under the new and improved method of dispersion went as far as five or ten would have gone under the old and wasteful casual method. Thus at last what had been a bunch of distinct coloured lilies grew to be a cuckoo-pint with an inclosing hood and a spike of minute central inconspicuous flowers.
[1] In all probability, the duckweed is not itself a really primitive type, but a degraded descendant of higher ancestors. This, however, does not prevent it from standing as an excellent representative of the real original unspecialised flowering plant, which must have been quite as simple in structure.
[2] Of course I do not mean to imply that daisies or primroses are descended from pinks; that would convey a wholly mistaken notion; but merely that the ancestors of the daisy once passed through a somewhat analogous stage.
[4] I owe my acknowledgments in much that follows to Mr. A. R. Wallace’s admirable work on Island Life.
[5] A lecture delivered at the London Institution, Finsbury Circus.
[6] The sedges are not, in all probability, a real natural family, but are a group of heterogeneous degraded lilies, containing almost all those kinds in which the reduced florets are covered by a single conspicuous glume-like bract. It will be seen from the sequel that these bracts are not truly homologous to the glumes or outer paleæ of grasses.
[8] All the potentillas have a double calyx, which certainly was not the case with the prime ancestor of the roses, or else the whole tribe would still retain it.
[9] Botanically and structurally these seeds, as we always call them, are really fruits; but the point is a purely technical one, with which it is quite unnecessary to bore the reader. I only mention it here to anticipate the sharp eyes of botanical critics.
[10] A lecture delivered at the Midland Institute, Birmingham.
[11] It is usual to treat these organs as staminodes—that is to say, abortive stamens. I know no reason for this classification, and the analogy of the scattered ovaries in the upper part of the Æthiopian lily leads me rather to regard them as altered pistils.
Hyphenation has been standardised.
page 52: Bricgstow changed to Brycgstow (Bristol)